

Mining of Ebola virus entry inhibitors identifies approved drugs as two-pore channel pore blockers
- PMID: 30408544
- PMCID: PMC7114365
- DOI: 10.1016/j.bbamcr.2018.10.022
Mining of Ebola virus entry inhibitors identifies approved drugs as two-pore channel pore blockers
Abstract
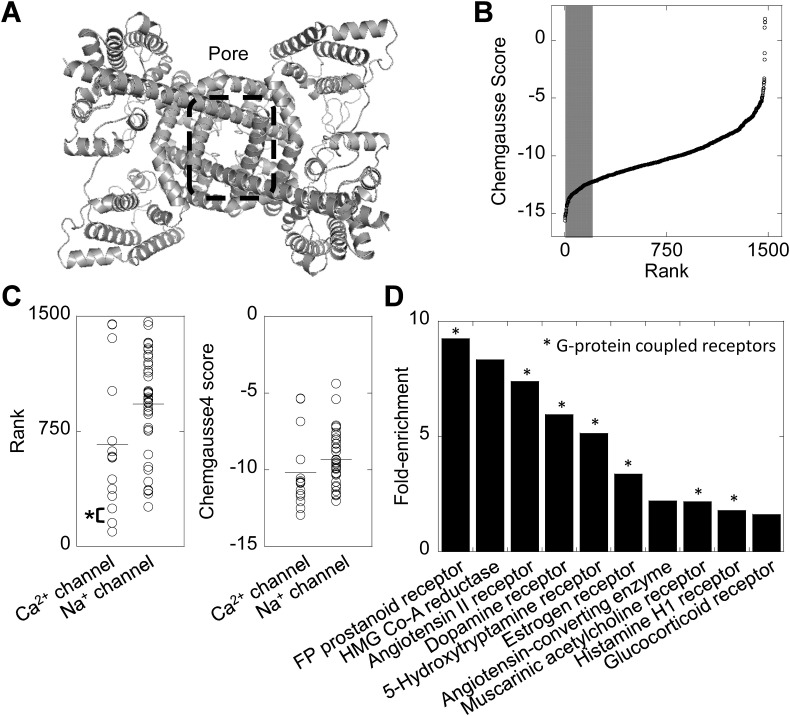
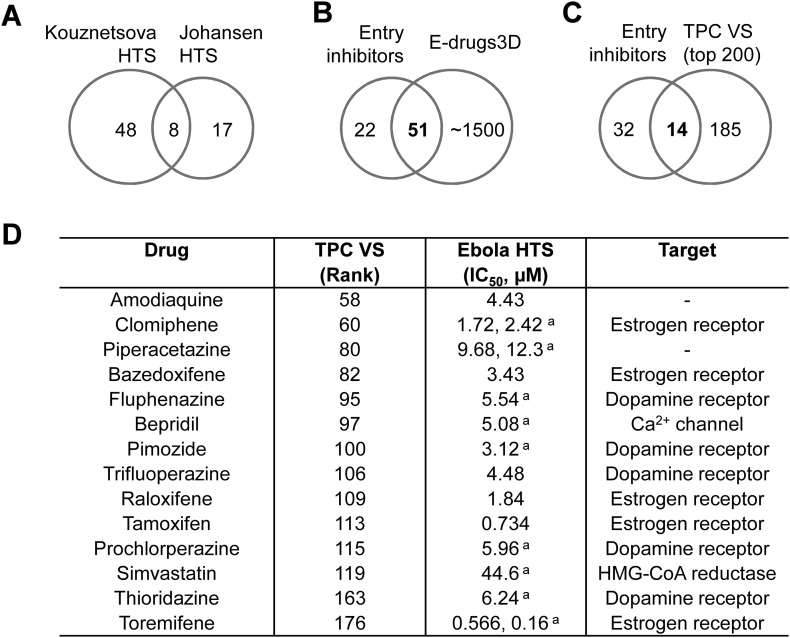
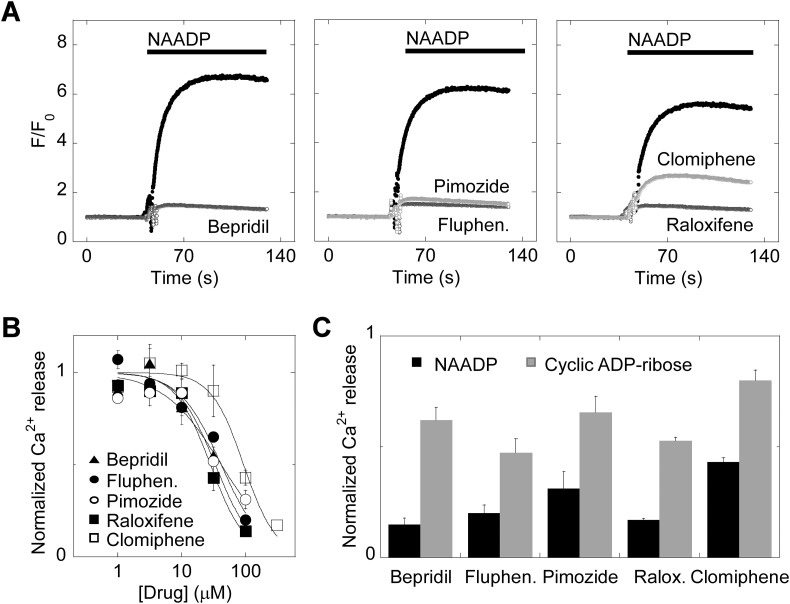
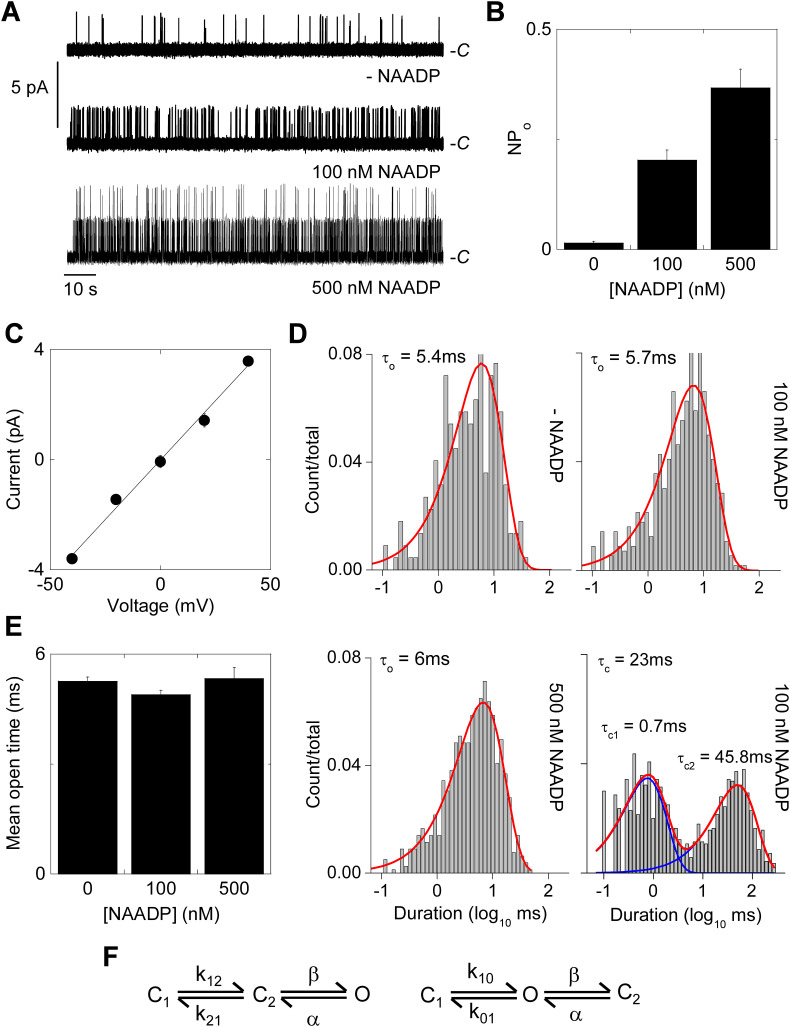
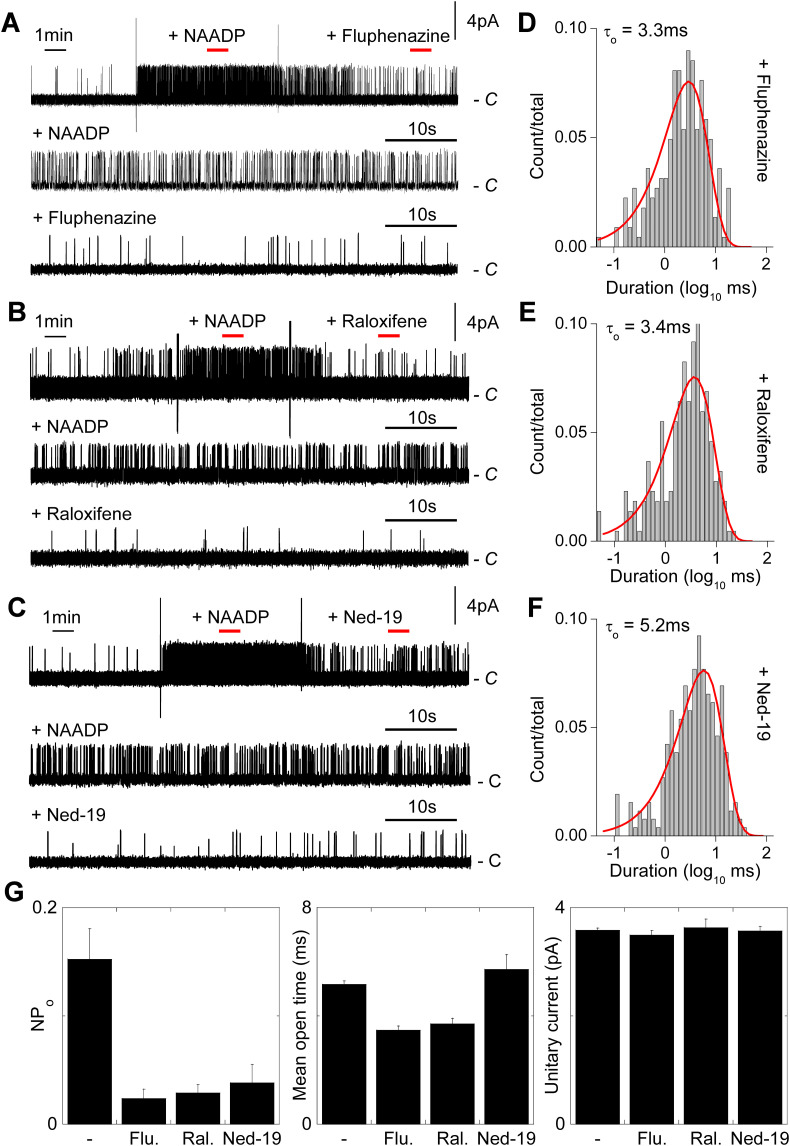
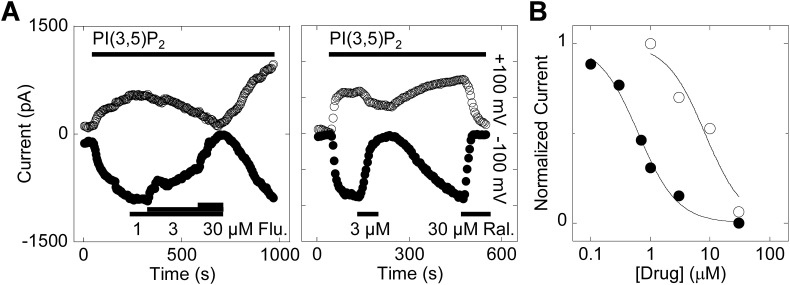
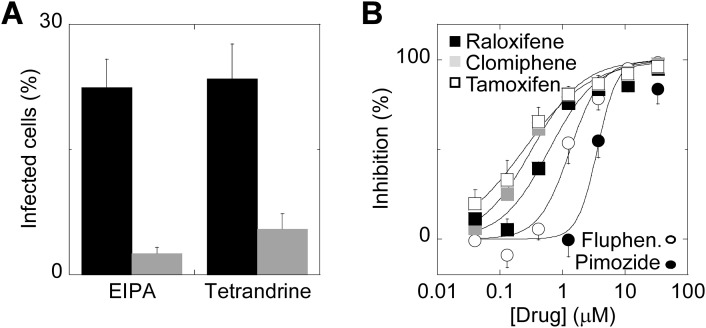
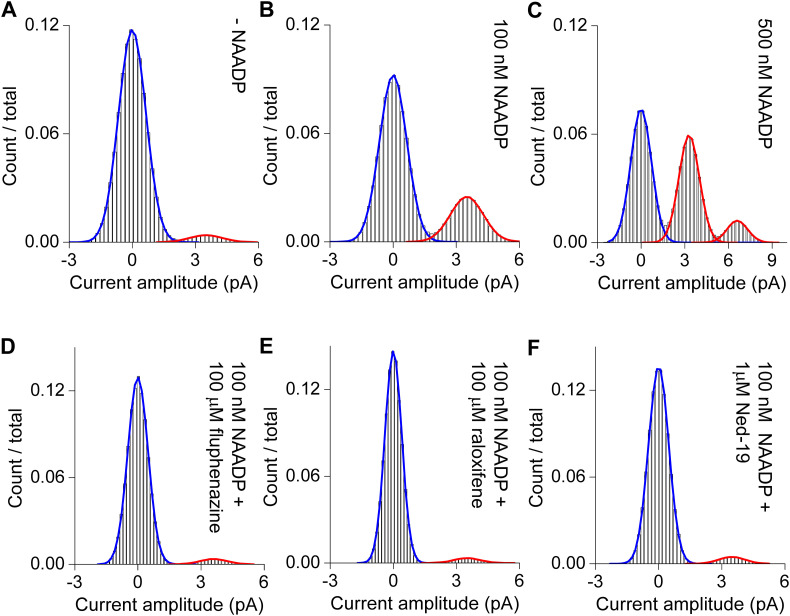
Two-pore channels (TPCs) are Ca2+-permeable ion channels localised to the endo-lysosomal system where they regulate trafficking of various cargoes including viruses. As a result, TPCs are emerging as important drug targets. However, their pharmacology is ill-defined. There are no approved drugs to target them. And their mechanism of ligand activation is largely unknown. Here, we identify a number of FDA-approved drugs as TPC pore blockers. Using a model of the pore of human TPC2 based on recent structures of mammalian TPCs, we virtually screened a database of ~1500 approved drugs. Because TPCs have recently emerged as novel host factors for Ebola virus entry, we reasoned that Ebola virus entry inhibitors may exert their effects through inhibition of TPCs. Cross-referencing hits from the TPC virtual screen with two recent high throughput anti-Ebola screens yielded approved drugs targeting dopamine and estrogen receptors as common hits. These compounds inhibited endogenous NAADP-evoked Ca2+ release from sea urchin egg homogenates, NAADP-mediated channel activity of TPC2 re-routed to the plasma membrane, and PI(3,5)P2-mediated channel activity of TPC2 expressed in enlarged lysosomes. Mechanistically, single channel analyses showed that the drugs reduced mean open time consistent with a direct action on the pore. Functionally, drug potency in blocking TPC2 activity correlated with inhibition of Ebola virus-like particle entry. Our results expand TPC pharmacology through the identification of approved drugs as novel blockers, support a role for TPCs in Ebola virus entry, and provide insight into the mechanisms underlying channel regulation. This article is part of a Special Issue entitled: ECS Meeting edited by Claus Heizmann, Joachim Krebs and Jacques Haiech.
Keywords: Ca(2+); Ebola virus; Lysosomes; NAADP; TPC2; Virtual screening.
Copyright © 2018. Published by Elsevier B.V.
Figures
References
-
- Churchill G.C., Okada Y., Thomas J.M., Genazzani A.A., Patel S., Galione A. NAADP mobilizes Ca2+ from reserve granules, lysosome-related organelles, in sea urchin eggs. Cell. 2002;111:703–708. - PubMed
-
- Lee H.C. NAADP-mediated calcium signaling. J. Biol. Chem. 2005;280(40):33693–33696. - PubMed
-
- Morgan A.J., Platt F.M., Lloyd-Evans E., Galione A. Molecular mechanisms of endolysosomal Ca2+ signalling in health and disease. Biochem. J. 2011;439:349–374. - PubMed
-
- Patel S., Cai X. Evolution of acid Ca2+ stores and their resident Ca2+-permeable channels. Cell Calcium. 2015;57:222–230. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- BB/G013721/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- MC_UU_00012/1/MRC_/Medical Research Council/United Kingdom
- MR/K015826/1/MRC_/Medical Research Council/United Kingdom
- MC_UU_12018/1/MRC_/Medical Research Council/United Kingdom
- H-1202/PUK_/Parkinson's UK/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
