

Gastrointestinal iron excretion and reversal of iron excess in a mouse model of inherited iron excess
- PMID: 30409795
- PMCID: PMC6442972
- DOI: 10.3324/haematol.2018.198382
Gastrointestinal iron excretion and reversal of iron excess in a mouse model of inherited iron excess
Abstract
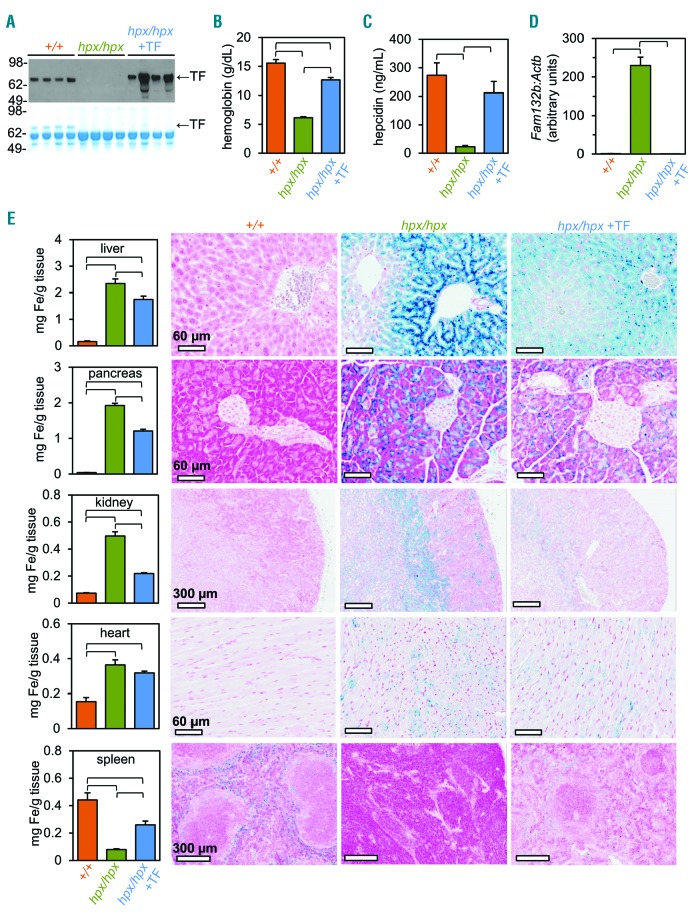
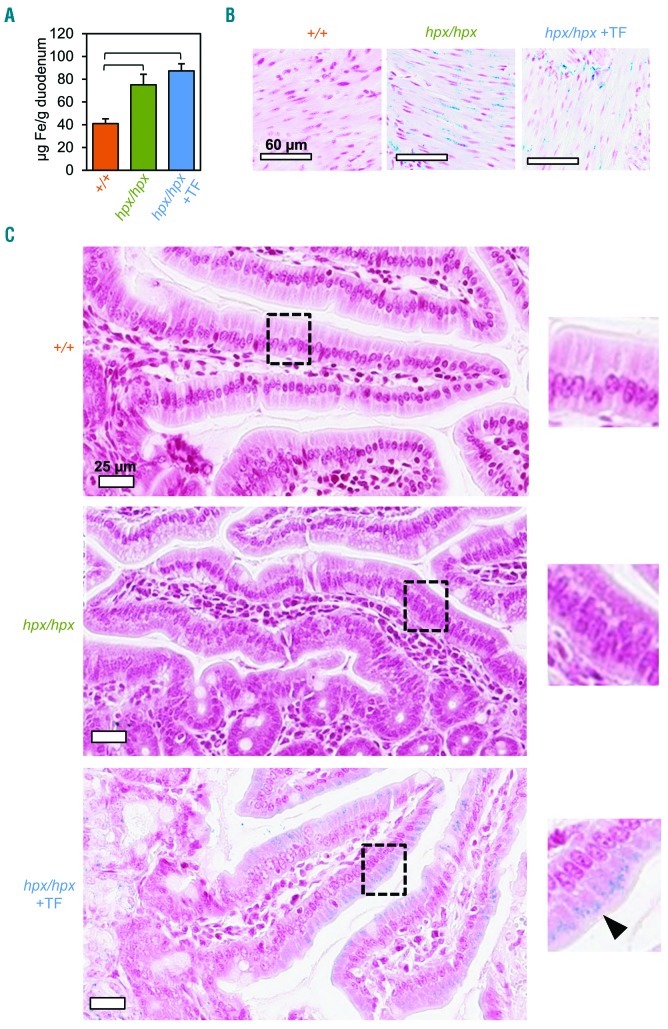
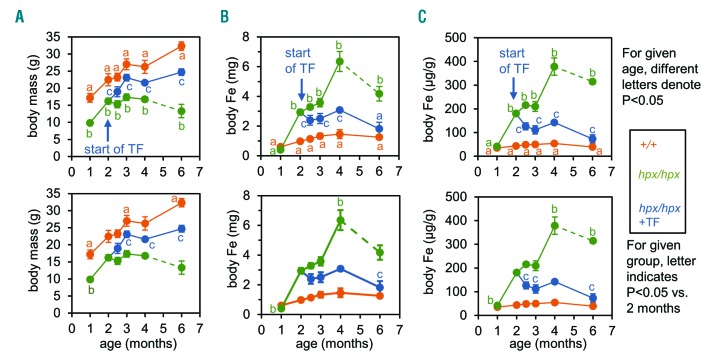
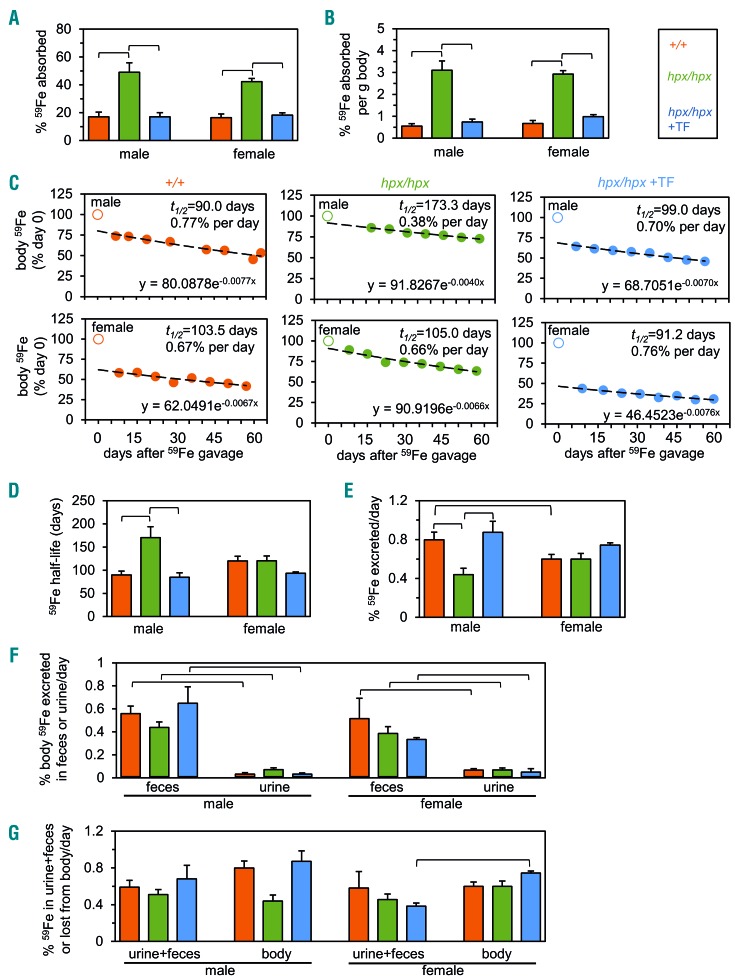
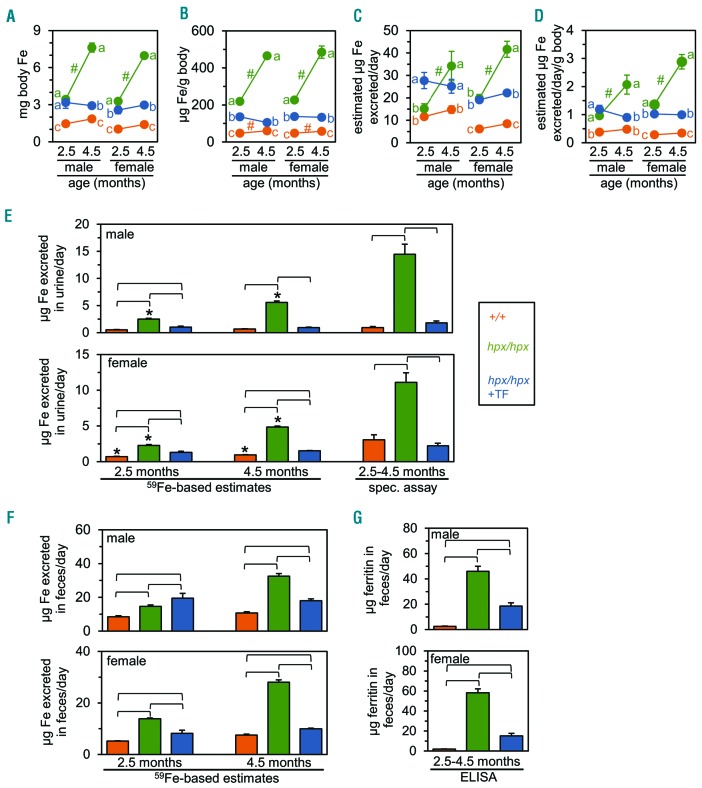
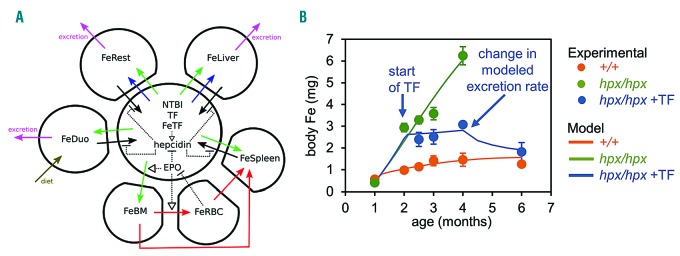
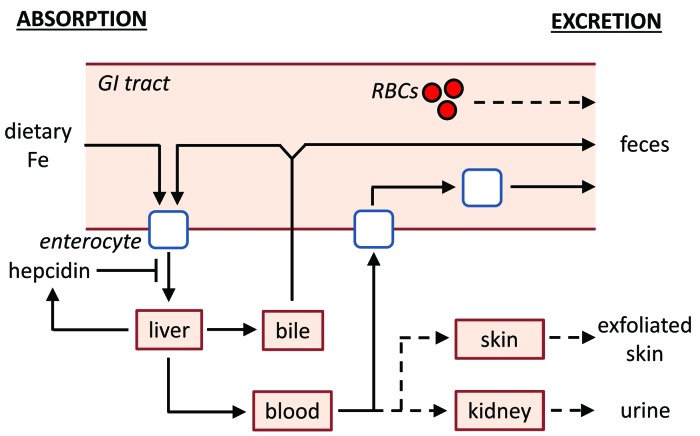
The current paradigm in the field of mammalian iron biology states that body iron levels are determined by dietary iron absorption, not by iron excretion. Iron absorption is a highly regulated process influenced by iron levels and other factors. Iron excretion is believed to occur at a basal rate irrespective of iron levels and is associated with processes such as turnover of intestinal epithelium, blood loss, and exfoliation of dead skin. Here we explore iron excretion in a mouse model of iron excess due to inherited transferrin deficiency. Iron excess in this model is attributed to impaired regulation of iron absorption leading to excessive dietary iron uptake. Pharmacological correction of transferrin deficiency not only normalized iron absorption rates and halted progression of iron excess but also reversed body iron excess. Transferrin treatment did not alter the half-life of 59Fe in mutant mice. 59Fe-based studies indicated that most iron was excreted via the gastrointestinal tract and suggested that iron-loaded mutant mice had increased rates of iron excretion. Direct measurement of urinary iron levels agreed with 59Fe-based predictions that urinary iron levels were increased in untreated mutant mice. Fecal ferritin levels were also increased in mutant mice relative to wild-type mice. Overall, these data suggest that mice have a significant capacity for iron excretion. We propose that further investigation into iron excretion is warranted in this and other models of perturbed iron homeostasis, as pharmacological targeting of iron excretion may represent a novel means of treatment for diseases of iron excess.
Copyright© 2019 Ferrata Storti Foundation.
Figures
References
-
- Papanikolaou G, Pantopoulos K. Systemic iron homeostasis and erythropoiesis. IUBMB Life. 2017;69(6):399–413. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
