

Random Neuronal Networks show homeostatic regulation of global activity while showing persistent changes in specific connectivity paths to theta burst stimuli
- PMID: 30410087
- PMCID: PMC6224599
- DOI: 10.1038/s41598-018-34634-x
Random Neuronal Networks show homeostatic regulation of global activity while showing persistent changes in specific connectivity paths to theta burst stimuli
Abstract
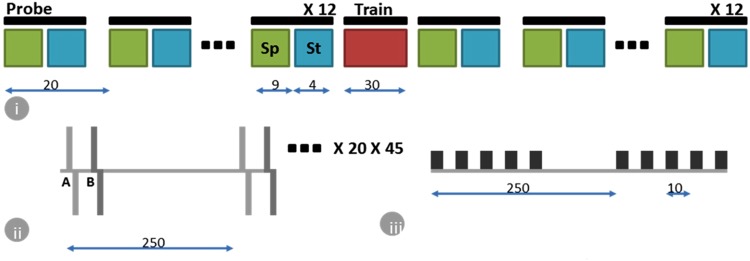
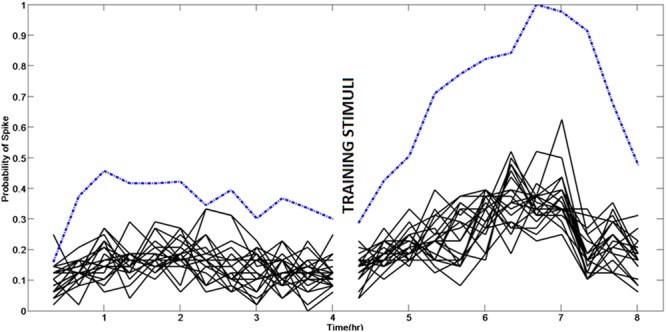
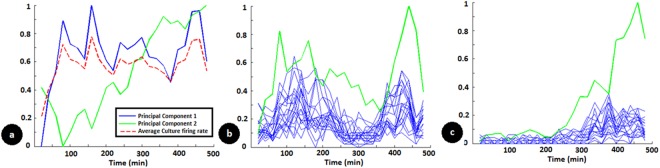
Learning in neuronal networks based on Hebbian principle has been shown to lead to destabilizing effects. Mechanisms have been identified that maintain homeostasis in such networks. However, the way in which these two opposing forces operate to support learning while maintaining stability is an active area of research. In this study, using neuronal networks grown on multi electrode arrays, we show that theta burst stimuli lead to persistent changes in functional connectivity along specific paths while the network maintains a global homeostasis. Simultaneous observations of spontaneous activity and stimulus evoked responses over several hours with theta burst training stimuli shows that global activity of the network quantified from spontaneous activity, which is disturbed due to theta burst stimuli is restored by homeostatic mechanisms while stimulus evoked changes in specific connectivity paths retain a memory trace of the training.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
