

A fast-evolving X-linked duplicate of importin-α2 is overexpressed in sex-ratio drive in Drosophila neotestacea
- PMID: 30411843
- PMCID: PMC6312747
- DOI: 10.1111/mec.14928
A fast-evolving X-linked duplicate of importin-α2 is overexpressed in sex-ratio drive in Drosophila neotestacea
Abstract
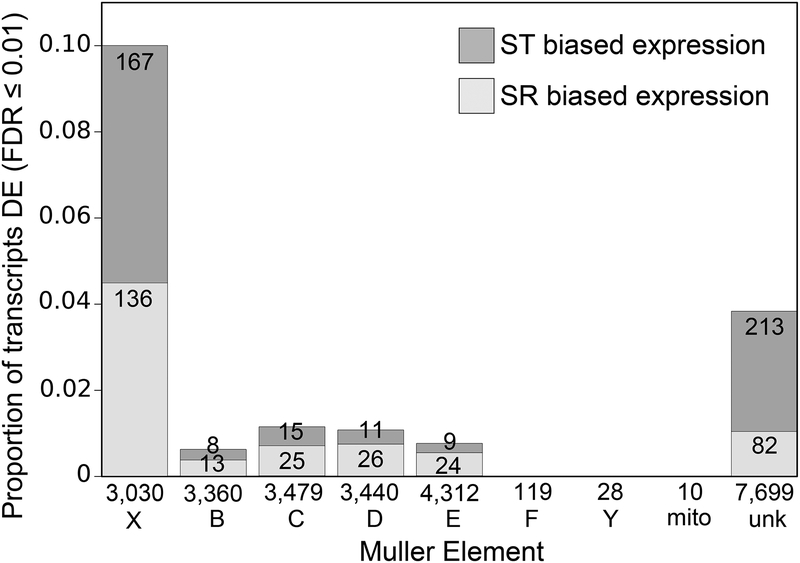
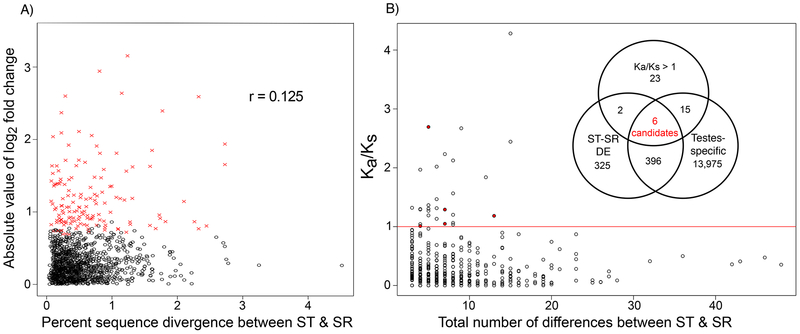
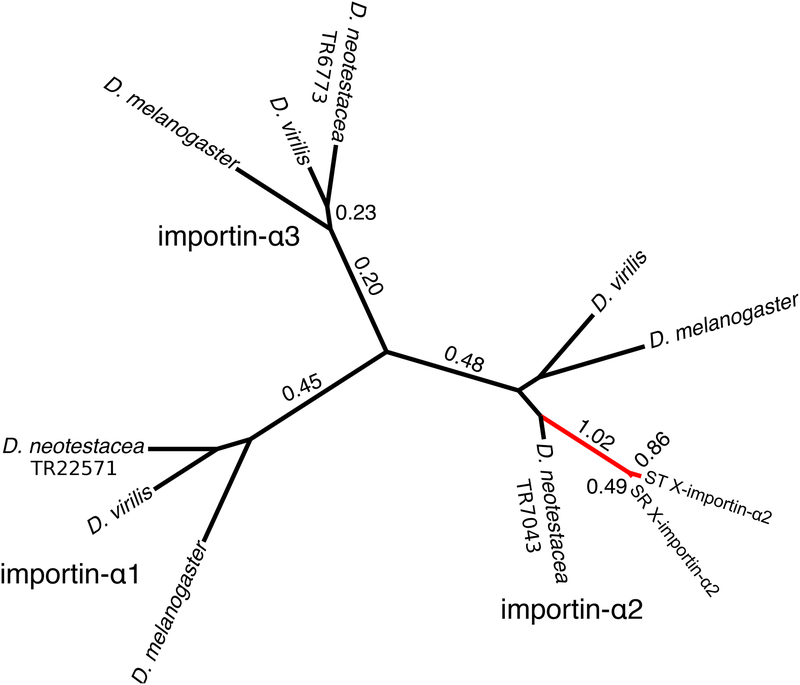
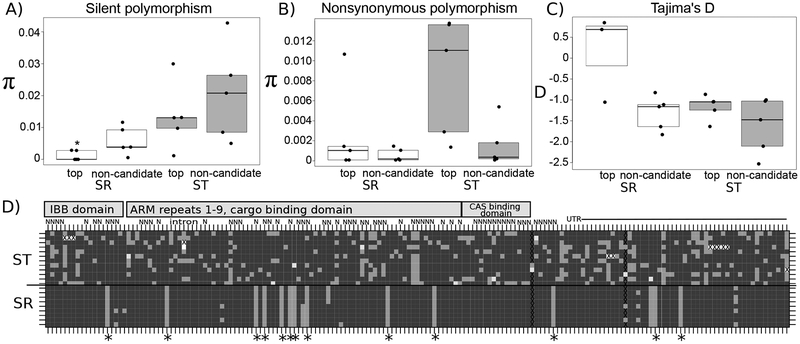
Selfish genetic elements that manipulate gametogenesis to achieve a transmission advantage are known as meiotic drivers. Sex-ratio X chromosomes (SR) are meiotic drivers that prevent the maturation of Y-bearing sperm in male carriers to result in the production of mainly female progeny. The spread of an SR chromosome can affect host genetic diversity and genome evolution, and can even cause host extinction if it reaches sufficiently high prevalence. Meiotic drivers have evolved independently many times, though only in a few cases is the underlying genetic mechanism known. In this study we use a combination of transcriptomics and population genetics to identify widespread expression differences between the standard (ST) and sex-ratio (SR) X chromosomes of the fly Drosophila neotestacea. We found the X chromosome is enriched for differentially expressed transcripts and that many of these X-linked differentially expressed transcripts had elevated Ka /Ks values between ST and SR, indicative of potential functional differences. We identified a set of candidate transcripts, including a testis-specific, X-linked duplicate of the nuclear transport gene importin-α2 that is overexpressed in SR. We find suggestions of positive selection in the lineage leading to the duplicate and that its molecular evolutionary patterns are consistent with relaxed purifying selection in ST. As these patterns are consistent with involvement in the mechanism of drive in this species, this duplicate is a strong candidate worthy of further functional investigation. Nuclear transport may be a common target for genetic conflict, as the mechanism of the autosomal Segregation Distorter drive system in D. melanogaster involves the same pathway.
Keywords: genetic conflict; meiotic drive; nuclear transport; spermatogenesis; testes.
© 2018 John Wiley & Sons Ltd.
Figures
References
-
- Anders S, & Huber W (2012). Differential expression of RNA-Seq data at the gene level–the DESeq package. Heidelberg, Germany: European Molecular Biology Laboratory (EMBL).
-
- Benjamini Y, & Hochberg Y (1995). Controlling the false discovery rate: a practical and powerful approach to multiple testing. Journal of the royal statistical society. Series B (Methodological), 289–300.
Publication types
MeSH terms
Substances
Grants and funding
- T32GM007103/GM/NIGMS NIH HHS/United States
- T32 GM007103/GM/NIGMS NIH HHS/United States
- Society for the Study of Evolution/International
- R00GM114714/NH/NIH HHS/United States
- 1457707/National Science Foundation/International
- University of Georgia/International
- 1149350/Division of Environmental Biology/International
- R00 GM114714/GM/NIGMS NIH HHS/United States
- 1457707/Division of Environmental Biology/International
- T32GM007103/NH/NIH HHS/United States
- 1149350/National Science Foundation/International
- R00GM114714/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
