

Interleukin-17-Producing γδ T Cells Originate from SOX13+ Progenitors that Are Independent of γδTCR Signaling
- PMID: 30413363
- PMCID: PMC6249057
- DOI: 10.1016/j.immuni.2018.09.010
Interleukin-17-Producing γδ T Cells Originate from SOX13+ Progenitors that Are Independent of γδTCR Signaling
Abstract
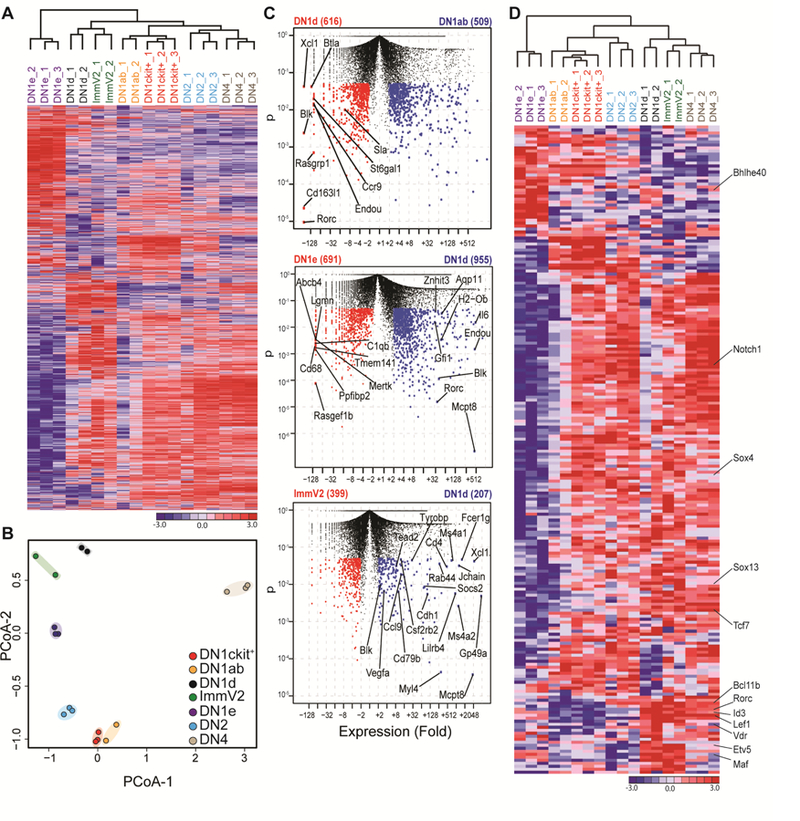
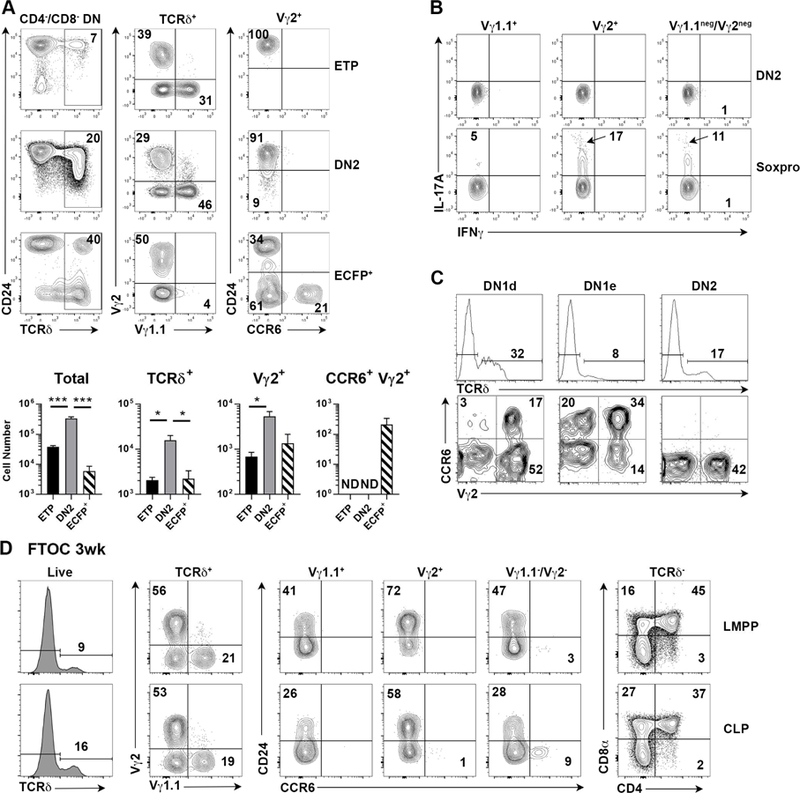
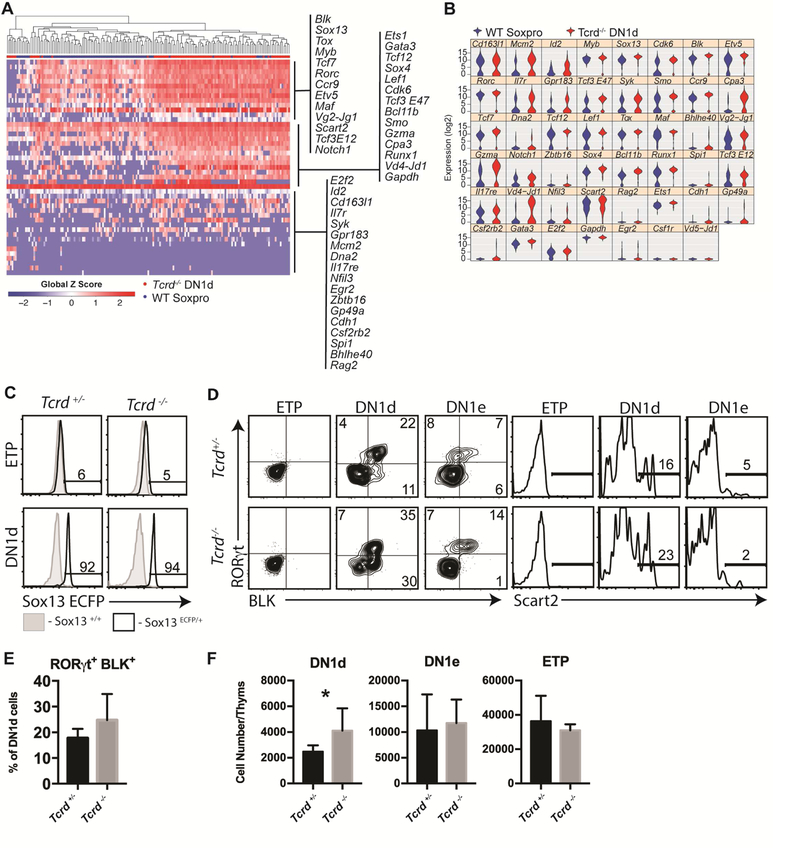
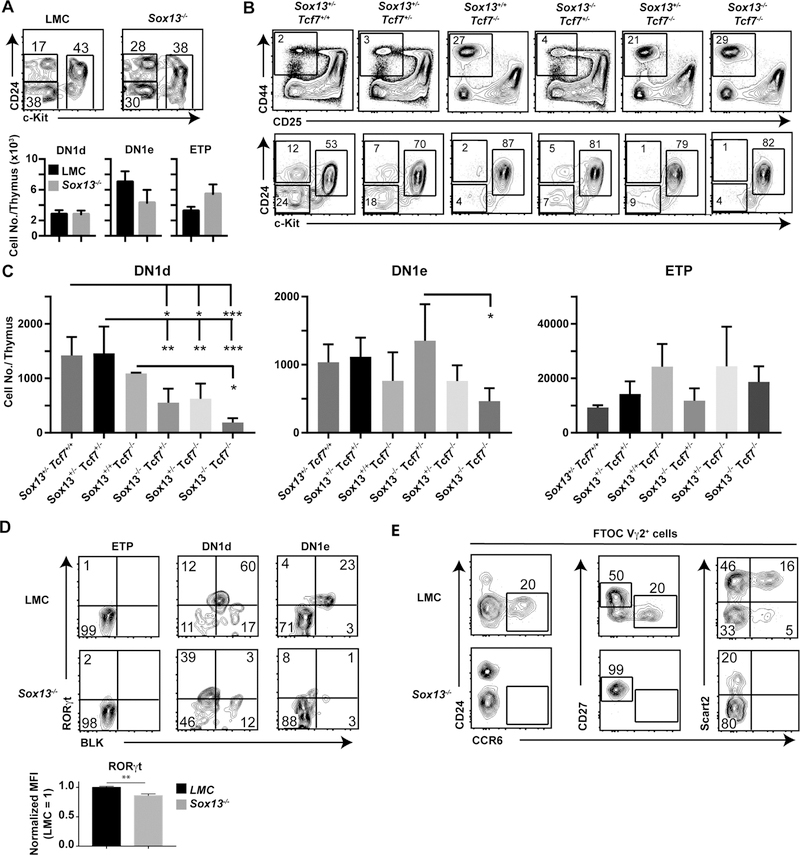
Lineage-committed αβ and γδ T cells are thought to originate from common intrathymic multipotent progenitors following instructive T cell receptor (TCR) signals. A subset of lymph node and mucosal Vγ2+ γδ T cells is programmed intrathymically to produce IL-17 (Tγδ17 cells), however the role of the γδTCR in development of these cells remains controversial. Here we generated reporter mice for the Tγδ17 lineage-defining transcription factor SOX13 and identified fetal-origin, intrathymic Sox13+ progenitors. In organ culture developmental assays, Tγδ17 cells derived primarily from Sox13+ progenitors, and not from other known lymphoid progenitors. Single cell transcriptome assays of the progenitors found in TCR-deficient mice demonstrated that Tγδ17 lineage programming was independent of γδTCR. Instead, generation of the lineage committed progenitors and Tγδ17 cells was controlled by TCF1 and SOX13. Thus, T lymphocyte lineage fate can be prewired cell-intrinsically and is not necessarily specified by clonal antigen receptor signals.
Keywords: IL-17; SOX13; T cell receptor; T cells; development; thymus; transcription factor.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests
The authors declare no competing interests.
Figures



References
-
- Boiers C, Carrelha J, Lutteropp M, Luc S, Green JC, Azzoni E, Woll PS, Mead AJ, Hultquist A,Swiers G, et al. (2013). Lymphomyeloid contribution of an immune-restricted progenitor emerging prior to definitive hematopoietic stem cells. Cell stem cell 13, 535–548. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
