

Genome-wide RNA structurome reprogramming by acute heat shock globally regulates mRNA abundance
- PMID: 30413617
- PMCID: PMC6275526
- DOI: 10.1073/pnas.1807988115
Genome-wide RNA structurome reprogramming by acute heat shock globally regulates mRNA abundance
Abstract
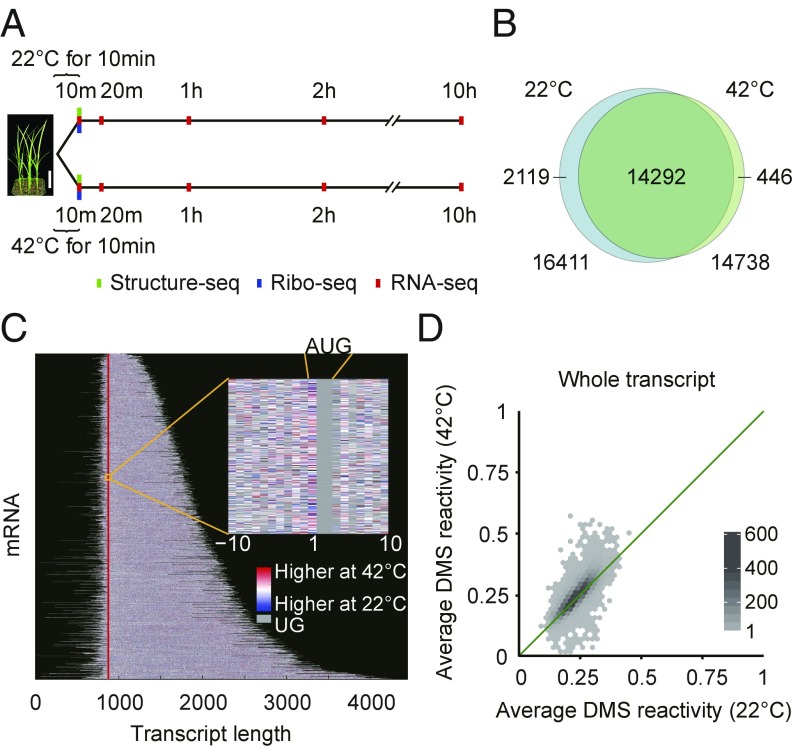
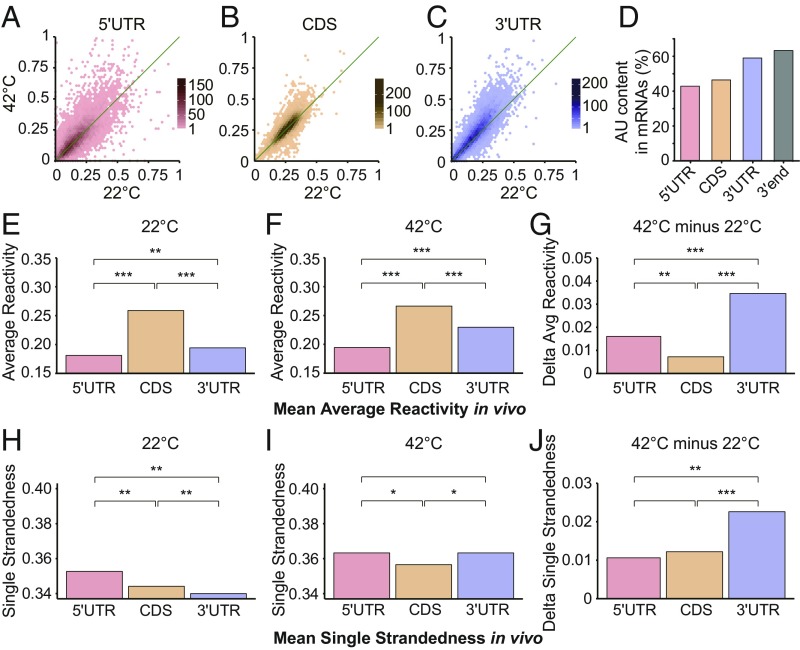
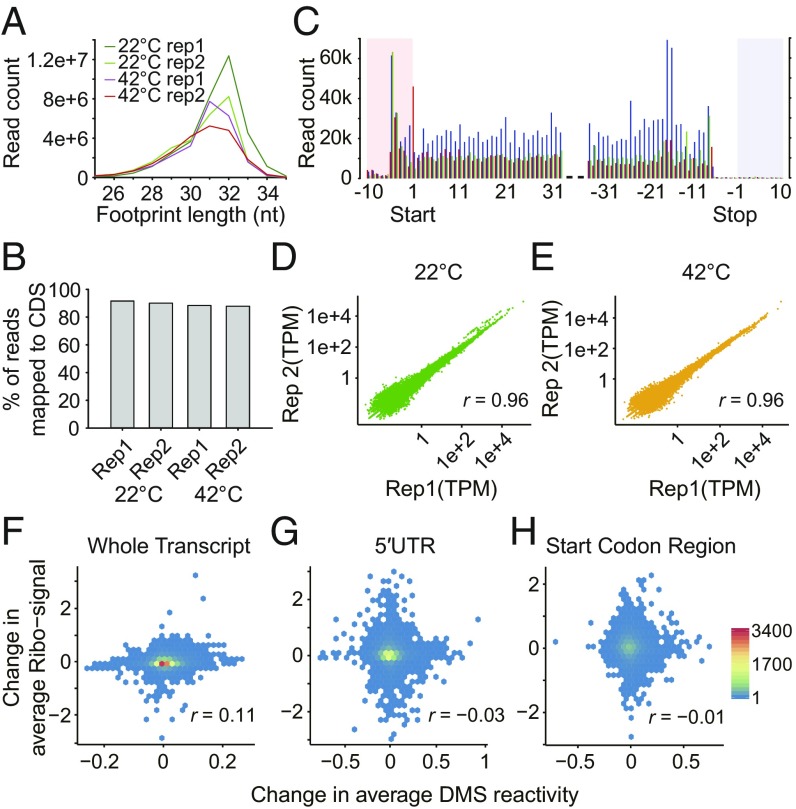
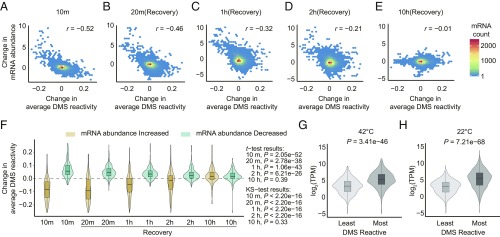
The heat shock response is crucial for organism survival in natural environments. RNA structure is known to influence numerous processes related to gene expression, but there have been few studies on the global RNA structurome as it prevails in vivo. Moreover, how heat shock rapidly affects RNA structure genome-wide in living systems remains unknown. We report here in vivo heat-regulated RNA structuromes. We applied Structure-seq chemical [dimethyl sulfate (DMS)] structure probing to rice (Oryza sativa L.) seedlings with and without 10 min of 42 °C heat shock and obtained structural data on >14,000 mRNAs. We show that RNA secondary structure broadly regulates gene expression in response to heat shock in this essential crop species. Our results indicate significant heat-induced elevation of DMS reactivity in the global transcriptome, revealing RNA unfolding over this biological temperature range. Our parallel Ribo-seq analysis provides no evidence for a correlation between RNA unfolding and heat-induced changes in translation, in contrast to the paradigm established in prokaryotes, wherein melting of RNA thermometers promotes translation. Instead, we find that heat-induced DMS reactivity increases correlate with significant decreases in transcript abundance, as quantified from an RNA-seq time course, indicating that mRNA unfolding promotes transcript degradation. The mechanistic basis for this outcome appears to be mRNA unfolding at both 5' and 3'-UTRs that facilitates access to the RNA degradation machinery. Our results thus reveal unexpected paradigms governing RNA structural changes and the eukaryotic RNA life cycle.
Keywords: RNA thermometer; RNA-seq; Structure-seq; heat shock; rice.
Copyright © 2018 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Battisti DS, Naylor RL. Historical warnings of future food insecurity with unprecedented seasonal heat. Science. 2009;323:240–244. - PubMed
-
- Kosová K, Vítámvás P, Prášil IT, Renaut J. Plant proteome changes under abiotic stress–Contribution of proteomics studies to understanding plant stress response. J Proteomics. 2011;74:1301–1322. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
