

Role of STIM1/ORAI1-mediated store-operated Ca2+ entry in skeletal muscle physiology and disease
- PMID: 30414508
- PMCID: PMC6290926
- DOI: 10.1016/j.ceca.2018.10.004
Role of STIM1/ORAI1-mediated store-operated Ca2+ entry in skeletal muscle physiology and disease
Abstract
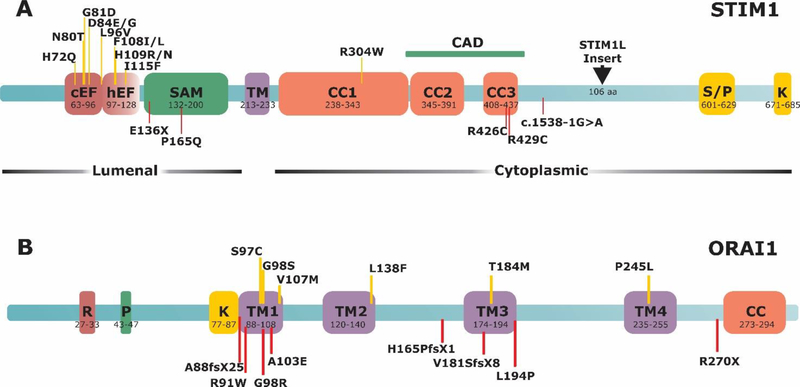
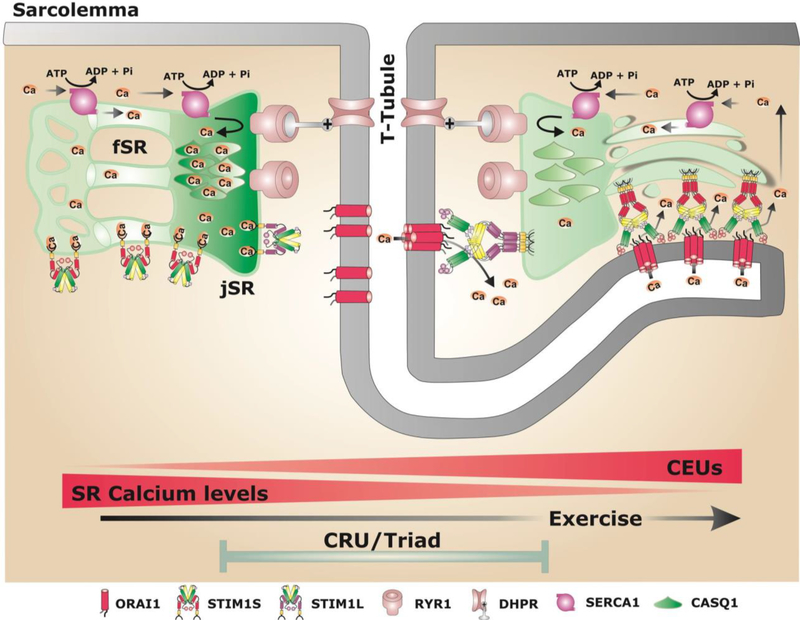
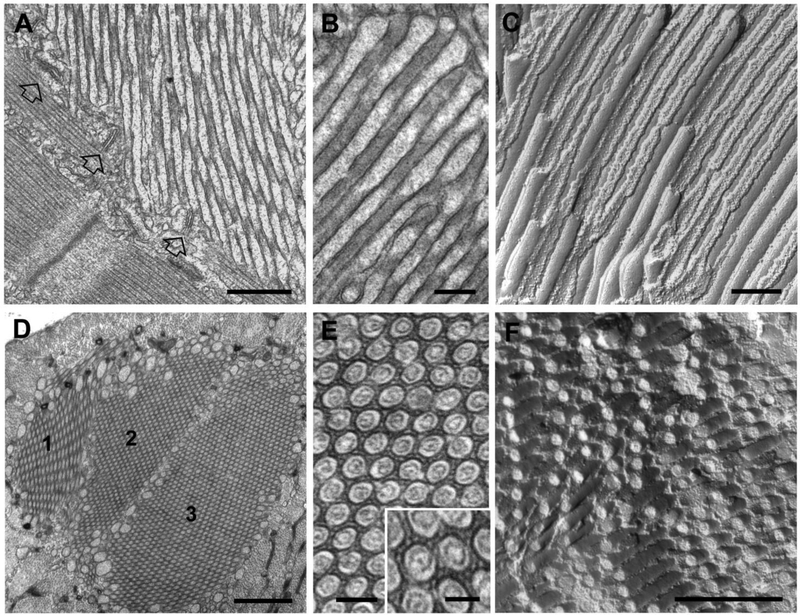
Store-operated Ca2+ entry (SOCE) is a Ca2+ entry mechanism activated by depletion of intracellular Ca2+ stores. In skeletal muscle, SOCE is mediated by an interaction between stromal-interacting molecule-1 (STIM1), the Ca2+ sensor of the sarcoplasmic reticulum, and ORAI1, the Ca2+-release-activated-Ca2+ (CRAC) channel located in the transverse tubule membrane. This review focuses on the molecular mechanisms and physiological role of SOCE in skeletal muscle, as well as how alterations in STIM1/ORAI1-mediated SOCE contribute to muscle disease. Recent evidence indicates that SOCE plays an important role in both muscle development/growth and fatigue. The importance of SOCE in muscle is further underscored by the discovery that loss- and gain-of-function mutations in STIM1 and ORAI1 result in an eclectic array of disorders with clinical myopathy as central defining component. Despite differences in clinical phenotype, all STIM1/ORAI1 gain-of-function mutations-linked myopathies are characterized by the abnormal accumulation of intracellular membranes, known as tubular aggregates. Finally, dysfunctional STIM1/ORAI1-mediated SOCE also contributes to the pathogenesis of muscular dystrophy, malignant hyperthermia, and sarcopenia. The picture to emerge is that tight regulation of STIM1/ORAI1-dependent Ca2+ signaling is critical for optimal skeletal muscle development/function such that either aberrant increases or decreases in SOCE activity result in muscle dysfunction.
Keywords: Ca(2+) signaling; Ca(2+)-release-activated-Ca(2+) (CRAC); Muscle fatigue; Tubular aggregate myopathy (TAM).
Copyright © 2018 Elsevier Ltd. All rights reserved.
Figures
References
-
- Putney JW Jr., A model for receptor-regulated calcium entry, Cell Calcium, 7 (1986) 1–12. - PubMed
-
- Hoth M, Penner R, Depletion of intracellular calcium stores activates a calcium current in mast cells, Nature, 355 (1992) 353–356. - PubMed
-
- Feske S, Gwack Y, Prakriya M, Srikanth S, Puppel SH, Tanasa B, Hogan PG, Lewis RS, Daly M, Rao A, A mutation in Orai1 causes immune deficiency by abrogating CRAC channel function, Nature, 441 (2006) 179–185. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
