

Spatial Organization of Single mRNPs at Different Stages of the Gene Expression Pathway
- PMID: 30415950
- PMCID: PMC6592633
- DOI: 10.1016/j.molcel.2018.10.010
Spatial Organization of Single mRNPs at Different Stages of the Gene Expression Pathway
Abstract
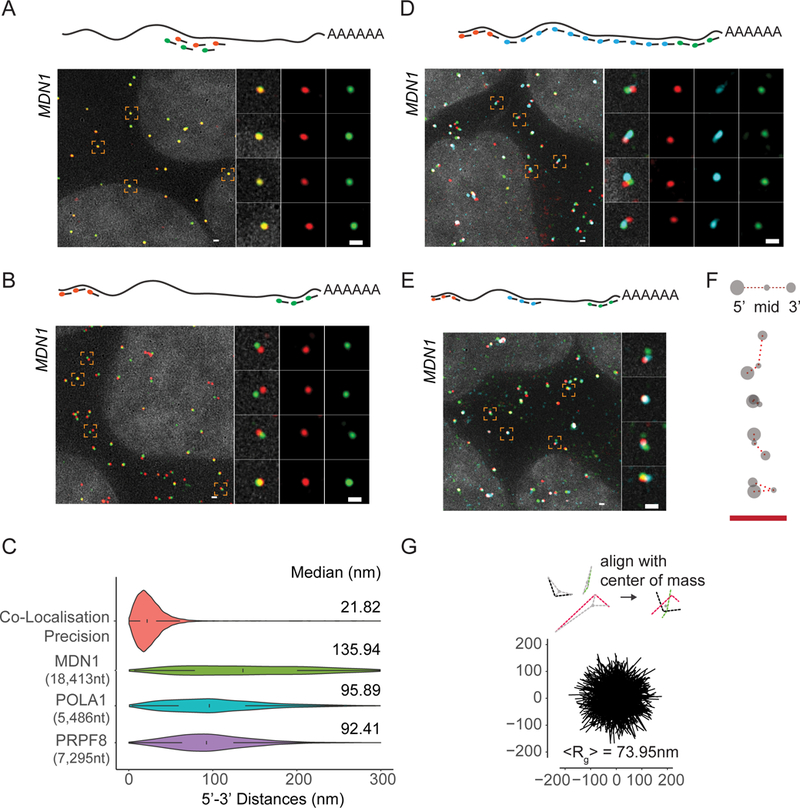
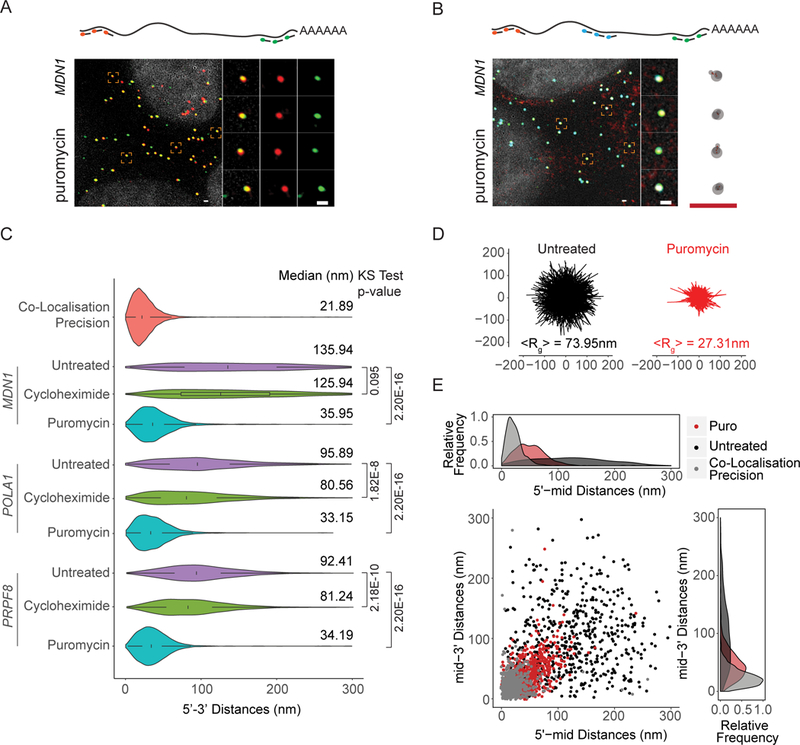
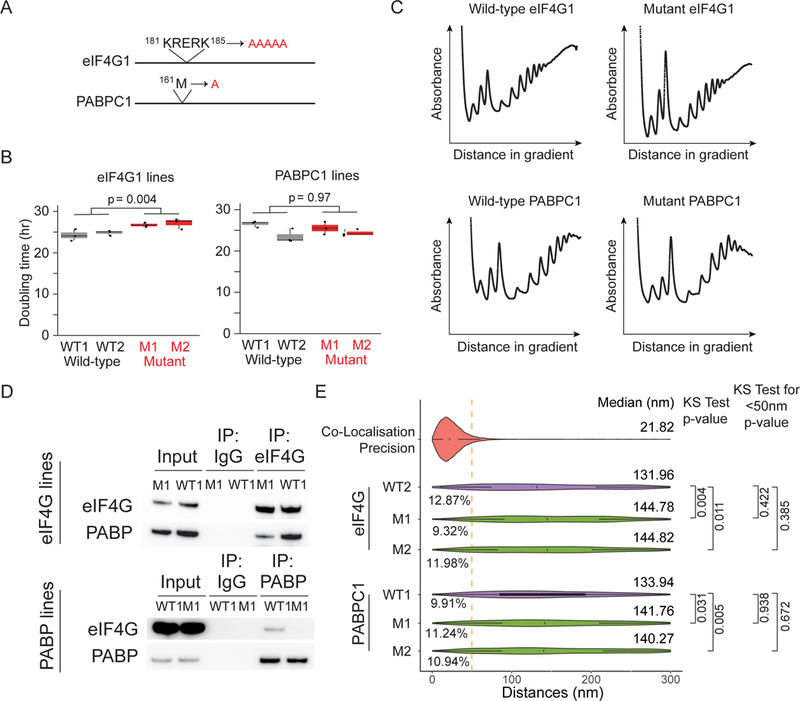
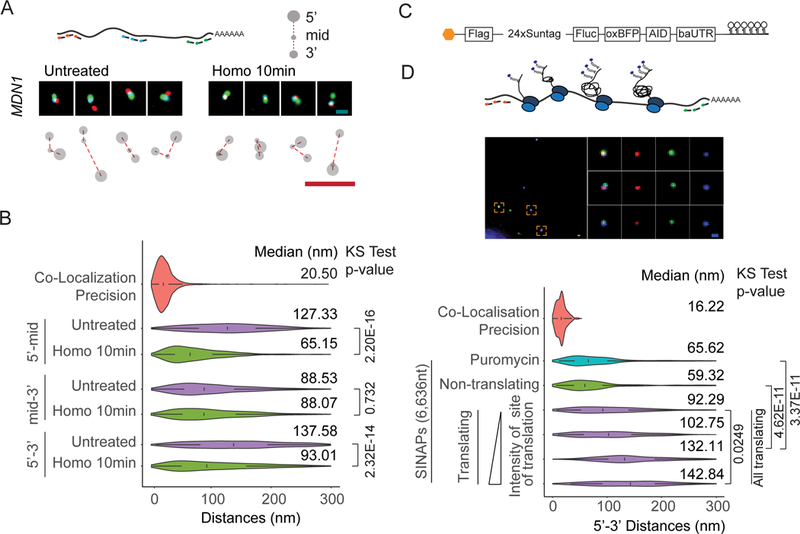
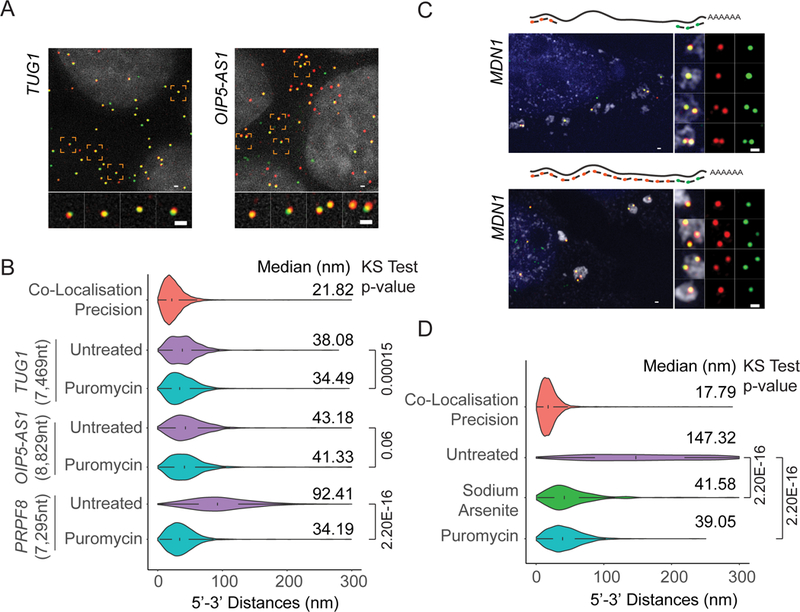
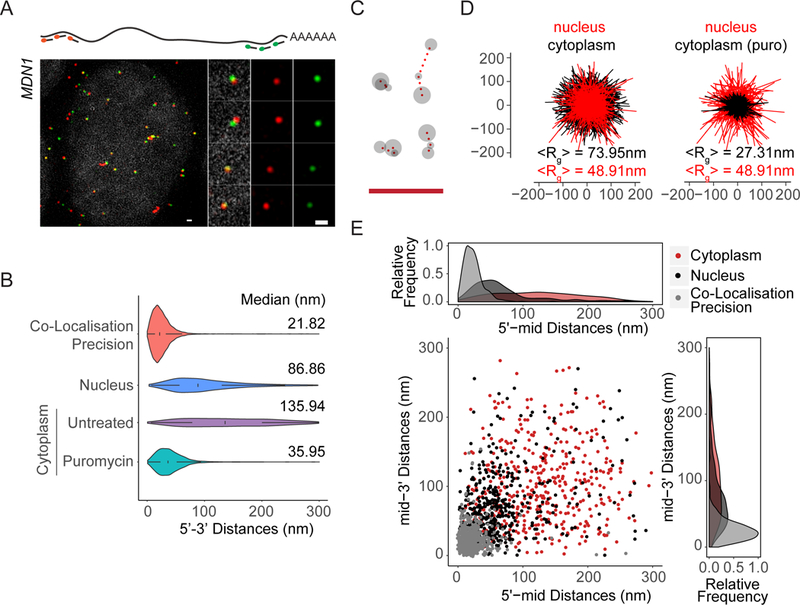
mRNAs form ribonucleoprotein complexes (mRNPs) by association with proteins that are crucial for mRNA metabolism. While the mRNP proteome has been well characterized, little is known about mRNP organization. Using a single-molecule approach, we show that mRNA conformation changes depending on its cellular localization and translational state. Compared to nuclear mRNPs and lncRNPs, association with ribosomes decompacts individual mRNAs, while pharmacologically dissociating ribosomes or sequestering them into stress granules leads to increased compaction. Moreover, translating mRNAs rarely show co-localized 5' and 3' ends, indicating either that mRNAs are not translated in a closed-loop configuration, or that mRNA circularization is transient, suggesting that a stable closed-loop conformation is not a universal state for all translating mRNAs.
Keywords: RNA compaction; RNA imaging; RNA structure; closed-loop translation; long non-coding RNAs; mRNP organization; smFISH; stress granules; super resolution microscopy; translation.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests
The authors declare no competing interests.
Figures
Comment in
-
Re-viewing the 3D Organization of mRNPs.Mol Cell. 2018 Nov 15;72(4):603-605. doi: 10.1016/j.molcel.2018.10.044. Mol Cell. 2018. PMID: 30444995
References
-
- Bhat M, Robichaud N, Hulea L, Sonenberg N, Pelletier J, Topisirovic I (2015). Targeting the translation machinery in cancer. Nat. Rev. Drug Discov 14, 261–278. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
