

Multiple Sclerosis-Associated Changes in the Composition and Immune Functions of Spore-Forming Bacteria
- PMID: 30417113
- PMCID: PMC6222044
- DOI: 10.1128/mSystems.00083-18
Multiple Sclerosis-Associated Changes in the Composition and Immune Functions of Spore-Forming Bacteria
Abstract
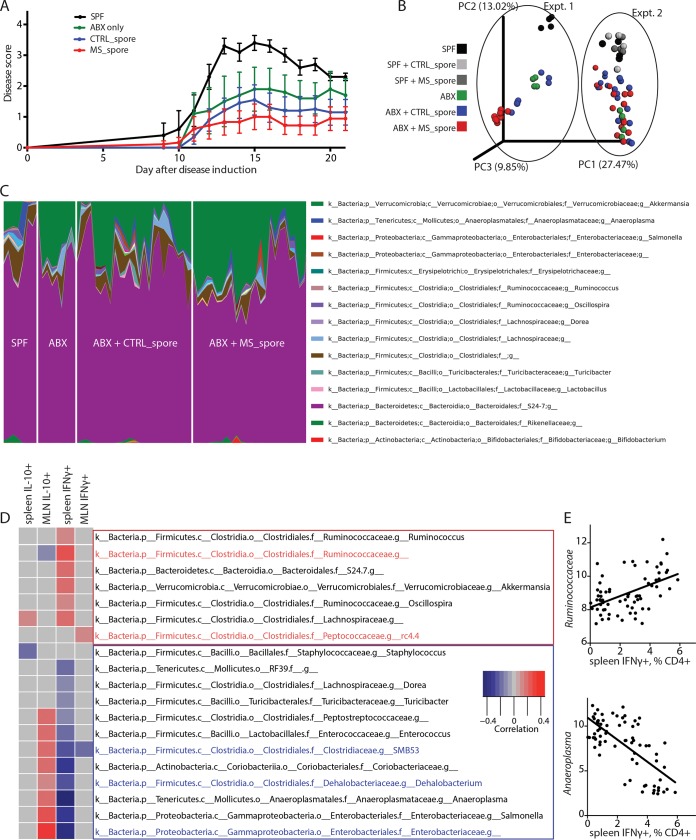
Multiple sclerosis (MS) is an autoimmune disease of the central nervous system characterized by adaptive and innate immune system dysregulation. Recent work has revealed moderate alteration of gut microbial communities in subjects with MS and in experimental, induced models. However, a mechanistic understanding linking the observed changes in the microbiota and the presence of the disease is still missing. Chloroform-resistant, spore-forming bacteria, which primarily belong to the classes Bacilli and Clostridia in the phylum Firmicutes, have been shown to exhibit immunomodulatory properties in vitro and in vivo, but they have not yet been characterized in the context of human disease. This study addresses the community composition and immune function of this bacterial fraction in MS. We identify MS-associated spore-forming taxa (primarily in the class Clostridia) and show that their presence correlates with impaired differentiation of IL-10-secreting, regulatory T lymphocytes in vitro. Colonization of antibiotic-treated mice with spore-forming bacteria allowed us to identify some bacterial taxa favoring IL-10+ lymphocyte differentiation and others inducing differentiation of proinflammatory, IFN-γ+ T lymphocytes. However, when fed into antibiotic-treated mice, both MS and control-derived spore-forming bacteria were able to induce similar IL-10-expressing Treg immunoregulatory responses, thus ameliorating symptoms of experimental allergic encephalomyelitis (EAE). Our analysis also identified Akkermansia muciniphila as a key organism that may interact either directly or indirectly with spore-forming bacteria to exacerbate the inflammatory effects of MS-associated gut microbiota. Thus, changes in the spore-forming fraction may influence T lymphocyte-mediated inflammation in MS. This experimental approach of isolating a subset of microbiota based on its functional characteristics may be useful to investigate other microbial fractions at greater depth. IMPORTANCE To address the impact of microbiome on disease development, it is essential to go beyond a descriptive study and evaluate the physiological importance of microbiome changes. Our study integrates computational analysis with in vitro and in vivo exploration of inflammatory properties of spore-forming microbial communities, revealing novel functional correlations. We specifically show that while small differences exist between the microbiomes of MS patients and healthy subjects, these differences are exacerbated in the chloroform-resistant fraction. We further demonstrate that, when purified from MS patients, this fraction is correlated with impaired immunomodulatory responses in vitro.
Keywords: immune mechanisms; multiple sclerosis; spore-forming bacteria.
Figures



References
-
- Gaboriau-Routhiau V, Rakotobe S, Lecuyer E, Mulder I, Lan A, Bridonneau C, Rochet V, Pisi A, De Paepe M, Brandi G, Eberl G, Snel J, Kelly D, Cerf-Bensussan N. 2009. The key role of segmented filamentous bacteria in the coordinated maturation of gut helper T cell responses. Immunity 31:677–689. doi:10.1016/j.immuni.2009.08.020. - DOI - PubMed
-
- Atarashi K, Tanoue T, Oshima K, Suda W, Nagano Y, Nishikawa H, Fukuda S, Saito T, Narushima S, Hase K, Kim S, Fritz JV, Wilmes P, Ueha S, Matsushima K, Ohno H, Olle B, Sakaguchi S, Taniguchi T, Morita H, Hattori M, Honda K. 2013. Treg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature 500:232–236. doi:10.1038/nature12331. - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
