

Visual physiology of the layer 4 cortical circuit in silico
- PMID: 30419013
- PMCID: PMC6258373
- DOI: 10.1371/journal.pcbi.1006535
Visual physiology of the layer 4 cortical circuit in silico
Abstract
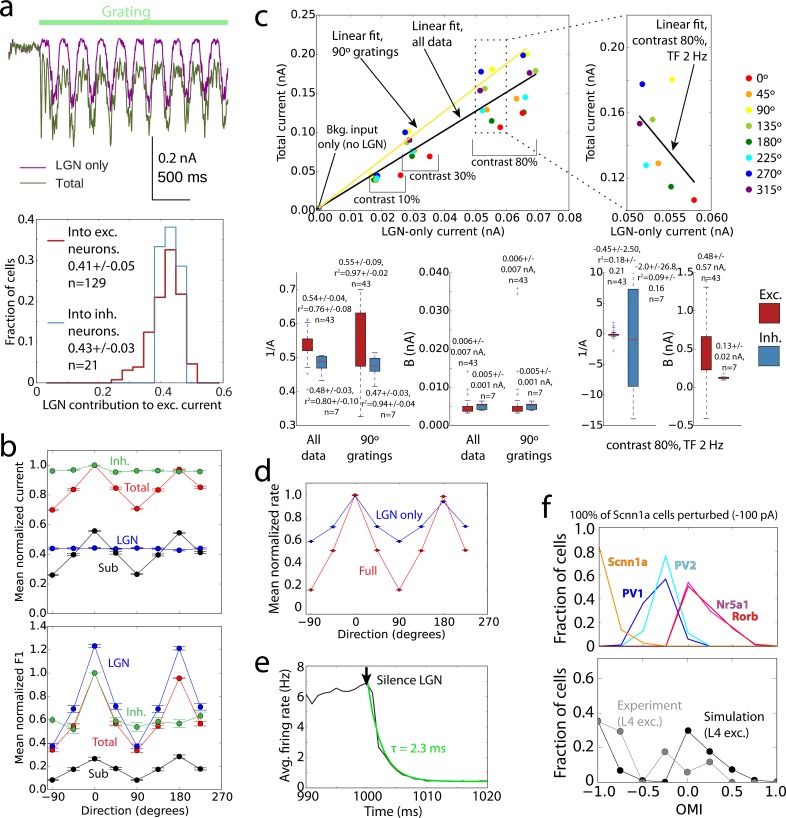
Despite advances in experimental techniques and accumulation of large datasets concerning the composition and properties of the cortex, quantitative modeling of cortical circuits under in-vivo-like conditions remains challenging. Here we report and publicly release a biophysically detailed circuit model of layer 4 in the mouse primary visual cortex, receiving thalamo-cortical visual inputs. The 45,000-neuron model was subjected to a battery of visual stimuli, and results were compared to published work and new in vivo experiments. Simulations reproduced a variety of observations, including effects of optogenetic perturbations. Critical to the agreement between responses in silico and in vivo were the rules of functional synaptic connectivity between neurons. Interestingly, after extreme simplification the model still performed satisfactorily on many measurements, although quantitative agreement with experiments suffered. These results emphasize the importance of functional rules of cortical wiring and enable a next generation of data-driven models of in vivo neural activity and computations.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures






References
-
- California Institute of Technology. Caltech Library Archives [Internet]. Available from: http://archives-dc.library.caltech.edu/islandora/object/ct1:483
-
- Markram H. et al. Reconstruction and Simulation of Neocortical Microcircuitry. Cell 163, 456–492 (2015). 10.1016/j.cell.2015.09.029 - DOI - PubMed
-
- Traub R.D. et al. Single-column thalamocortical network model exhibiting gamma oscillations, sleep spindles, and epileptogenic bursts. J. Neurophysiol. 93, 2194–2232 (2005). 10.1152/jn.00983.2004 - DOI - PubMed
-
- Zhu W, Shelley M, and Shapley R. A neuronal network model of primary visual cortex explains spatial frequency selectivity. J. Comput. Neurosci. 26, 271–287 (2009). 10.1007/s10827-008-0110-x - DOI - PubMed
-
- Buzsaki G., and Mizuseki K. The log-dynamic brain: how skewed distributions affect network operations. Nat Rev Neurosci. 15, 264–278 (2014). 10.1038/nrn3687 - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
