

Cholesterol intake and statin use regulate neuronal G protein-gated inwardly rectifying potassium channels
- PMID: 30420402
- PMCID: PMC6314255
- DOI: 10.1194/jlr.M081240
Cholesterol intake and statin use regulate neuronal G protein-gated inwardly rectifying potassium channels
Abstract
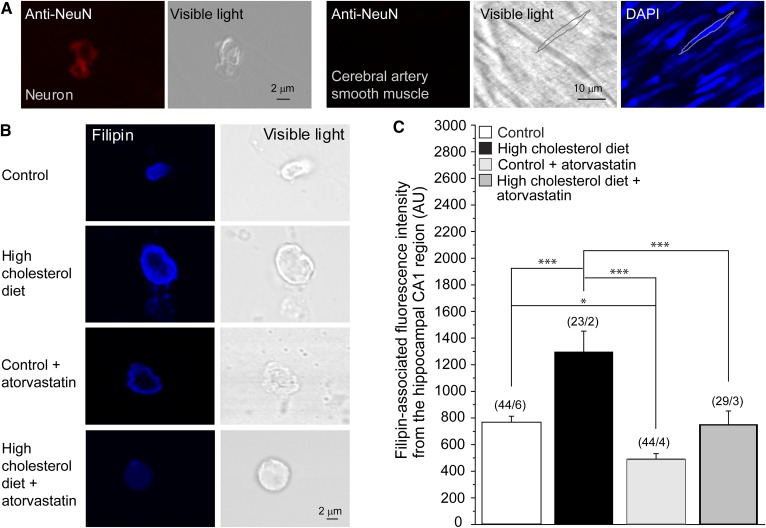
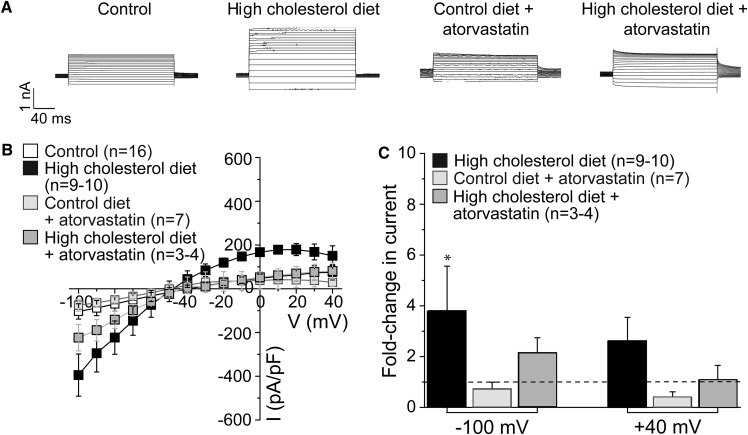
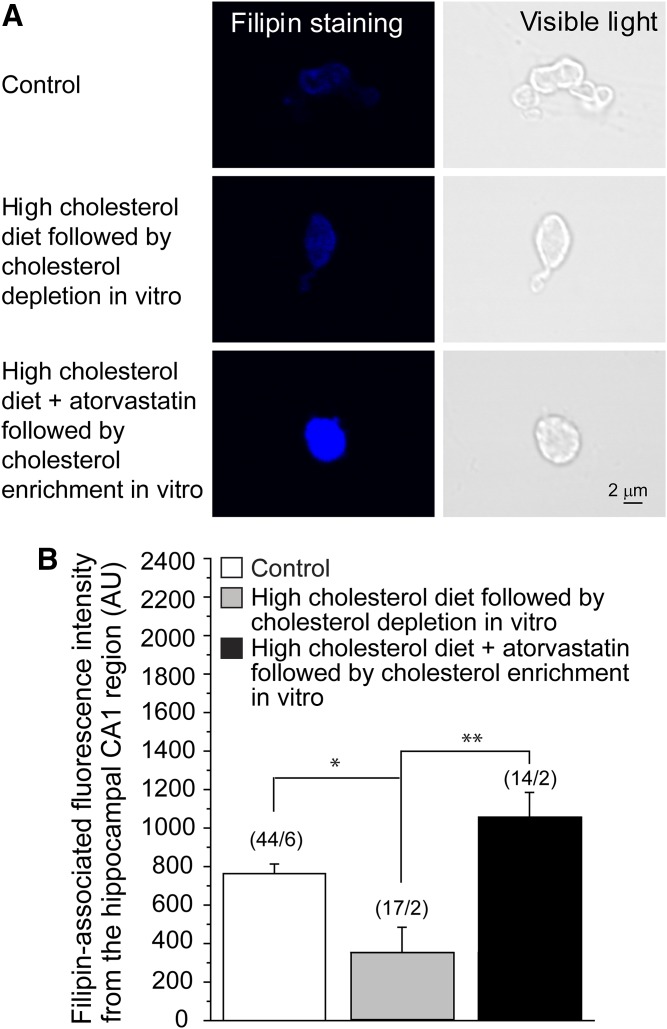
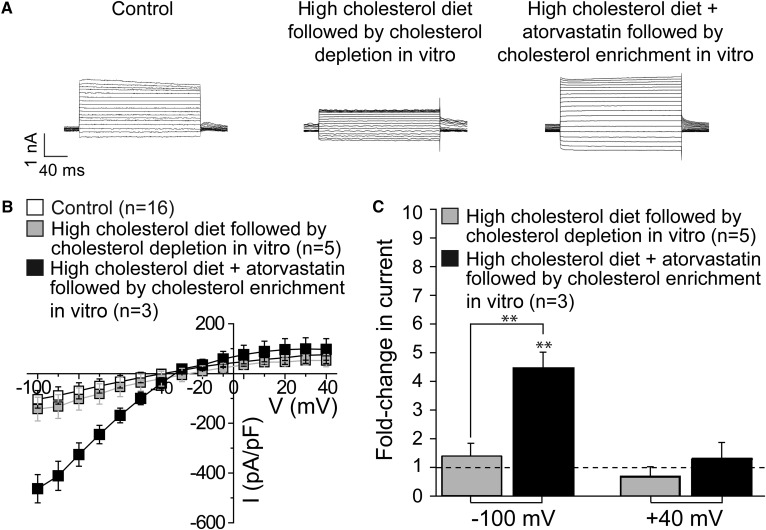
Cholesterol, a critical component of the cellular plasma membrane, is essential for normal neuronal function. Cholesterol content is highest in the brain, where most cholesterol is synthesized de novo; HMG-CoA reductase controls the synthesis rate. Despite strict control, elevated blood cholesterol levels are common and are associated with various neurological disorders. G protein-gated inwardly rectifying potassium (GIRK) channels mediate the actions of inhibitory brain neurotransmitters. Loss of GIRK function enhances neuron excitability; gain of function reduces neuronal activity. However, the effect of dietary cholesterol or HMG-CoA reductase inhibition (i.e., statin therapy) on GIRK function remains unknown. Using a rat model, we compared the effects of a high-cholesterol versus normal diet both with and without atorvastatin, a widely prescribed HMG-CoA reductase inhibitor, on neuronal GIRK currents. The high-cholesterol diet increased hippocampal CA1 region cholesterol levels and correspondingly increased neuronal GIRK currents. Both phenomena were reversed by cholesterol depletion in vitro. Atorvastatin countered the high-cholesterol diet effects on neuronal cholesterol content and GIRK currents; these effects were reversed by cholesterol enrichment in vitro. Our findings suggest that high-cholesterol diet and atorvastatin therapy affect ion channel function in the brain by modulating neuronal cholesterol levels.
Keywords: 3-hydroxy-3-methylglutaryl-CoA reductase; CA1 hippocampal neuron; brain lipids; dietary cholesterol; inwardly rectifying potassium channel; lipid mediators; lipid signaling.
Figures

References
-
- Yeagle P. L. 1985. Cholesterol and the cell membrane. Biochim. Biophys. Acta. 822: 267–287. - PubMed
-
- Yeagle P. L. 1991. Modulation of membrane function by cholesterol. Biochimie. 73: 1303–1310. - PubMed
-
- Rosenhouse-Dantsker A., Mehta D., and Levitan I.. 2012. Regulation of ion channels by membrane lipids. Compr. Physiol. 2: 31–68. - PubMed
-
- Ramprasad O. G., Srinivas G., Rao K. S., Joshi P., Thiery J. P., Dufour S., and Pande G.. 2007. Changes in cholesterol levels in the plasma membrane modulate cell signaling and regulate cell adhesion and migration on fibronectin. Cell Motil. Cytoskeleton. 64: 199–216. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
