

Shelterin and subtelomeric DNA sequences control nucleosome maintenance and genome stability
- PMID: 30420521
- PMCID: PMC6322387
- DOI: 10.15252/embr.201847181
Shelterin and subtelomeric DNA sequences control nucleosome maintenance and genome stability
Abstract
Telomeres and the shelterin complex cap and protect the ends of chromosomes. Telomeres are flanked by the subtelomeric sequences that have also been implicated in telomere regulation, although their role is not well defined. Here, we show that, in Schizosaccharomyces pombe, the telomere-associated sequences (TAS) present on most subtelomeres are hyper-recombinogenic, have metastable nucleosomes, and unusual low levels of H3K9 methylation. Ccq1, a subunit of shelterin, protects TAS from nucleosome loss by recruiting the heterochromatic repressor complexes CLRC and SHREC, thereby linking nucleosome stability to gene silencing. Nucleosome instability at TAS is independent of telomeric repeats and can be transmitted to an intrachromosomal locus containing an ectopic TAS fragment, indicating that this is an intrinsic property of the underlying DNA sequence. When telomerase recruitment is compromised in cells lacking Ccq1, DNA sequences present in the TAS promote recombination between chromosomal ends, independent of nucleosome abundance, implying an active function of these sequences in telomere maintenance. We propose that Ccq1 and fragile subtelomeres co-evolved to regulate telomere plasticity by controlling nucleosome occupancy and genome stability.
Keywords: genome stability; heterochromatin; nucleosomes; shelterin; telomeres.
© 2018 The Authors.
Figures
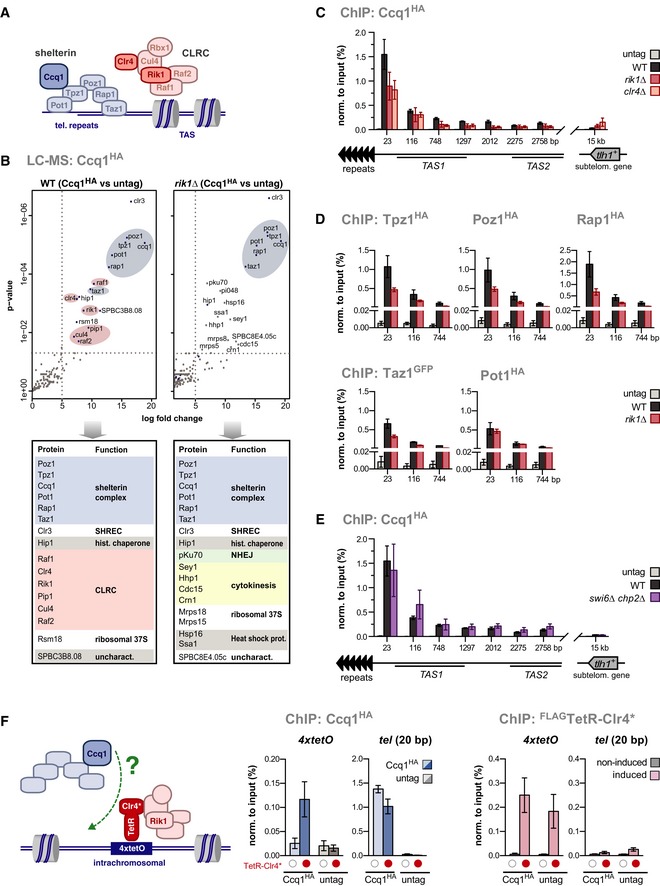
- A
Scheme of the shelterin complex and CLRC at telomeres.
- B
Mass spectrometry analysis of proteins co‐purified with Ccq1HA. Shown is a volcano plot for proteins significantly enriched in Ccq1HA relative to untagged Ccq1 (left panel: WT cells; right panel: rik1∆ cells). Members of the CLRC complex and shelterin are highlighted in red and blue, respectively. Bottom panel displays proteins enriched (log2 ≥ 5 or P ≤ 0.01).
- C–E
ChIP‐qPCR analysis of epitope‐tagged shelterin components in WT and mutant strains as indicated (negative control: untagged strain). Positions on x‐axis denote distances relative to telomeric repeats (telomere‐proximal PCR primer) (n = 3 independent experiments).
- F
ChIP analysis of Ccq1HA and FLAG‐TetR‐Clr4* (see scheme). Expression and tethering of FLAG‐TetR‐Clr4* are controlled by thiamine/AHT addition (red dots indicate induced/tethered FLAG‐TetR‐Clr4*; empty circles indicate non‐induced/non‐bound controls) (n = 3 independent experiments).
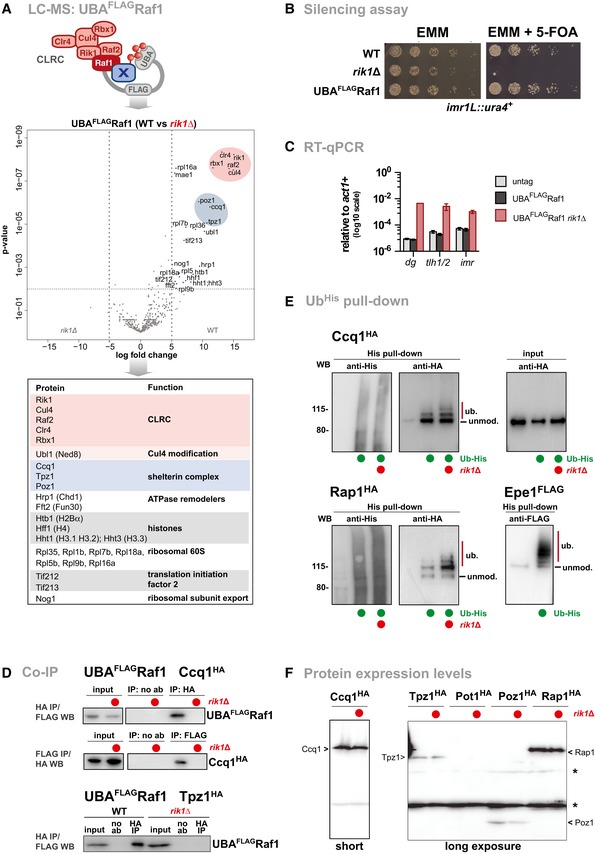
Mass spectrometry analysis of proteins co‐purified with Raf1 fused to the ubiquitin‐associated (UBA) domain of the ubiquitin receptor Dsk2 from S. cerevisiae (UBAFLAGRaf1). This method is based on the Ubiquitin Ligase Trapping method originally developed in S. cerevisiae 57 , which stabilizes the binding of substrates to the ubiquitin ligase. The volcano plot depicts proteins enriched in UBAFLAGRaf1 WT cells relative to UBAFLAGRaf1 rik1Δ. Members of the CLRC and shelterin complexes are highlighted in red and blue, respectively. Bottom panel displays nuclear proteins enriched (log2 ≥ 5 or P ≤ 0.01).
Silencing reporter assay at imr1L::ura4 +. Fivefold serial dilution on non‐selective (EMM) and selective media (EMM + 5‐FOA) of WT, rik∆, and UBAFLAGRaf1 cells. The N‐terminal fusion does not interfere with the function of CLRC in heterochromatin formation. The rik1∆ strain was used as negative control.
RT–qPCR analysis in strains expressing untagged Raf1, UBAFLAGRaf1, and UBAFLAGRaf1 rik1∆ (n = 3 independent experiments). Data are normalized to input and represented as mean ± SEM.
Co‐immunoprecipitation of Raf1 with Ccq1 (top and middle panel) and Tpz1 (bottom panel) in presence and absence of Rik1. Strains expressing endogenous levels of UBAFLAGRaf1 and Ccq1HA or Tpz1HA were subjected to anti‐HA immunoprecipitation in the presence of Benzonase (indicating that the interactions are independent of DNA or RNA; negative control: no antibody, noAb). Input and immunoprecipitated material were analyzed by anti‐FLAG immunoblots. For Ccq1HA, the reciprocal experiment is also shown.
Ubiquitin pull‐down experiments. Top panels shows in vivo ubiquitylation of Ccq1HA in WT and rik1∆ cells expressing 6His‐ubiquitin. Left panels: precipitated 6His‐ubiquitin conjugates (20%) analyzed by anti‐His and anti‐HA immunoblots. Right panel: input fraction (0.1%) analyzed by anti‐HA immunoblot. The bottom panels show 6His‐ubiquitin pull‐down assays for Rap1HA in WT and rik1∆ cells (left) and Epe1FLAG in WT cells (right). Epe1 is shown as an example for poly‐ubiquitylation. WT cells expressing untagged ubiquitin are used as negative control.
Anti‐HA immunoblot of epitope‐tagged shelterin proteins expressed from their endogenous locus in WT and rik1∆ cells. The asterisks denote unspecific signals derived from cross‐reactions with the anti‐HA antibody. Note that Ccq1 is expressed at higher levels compared to other shelterin subunits; endogenous levels of Pot1 are low and not detected by the immunoblot. Estimated sizes (kDa) of the tagged proteins: Ccq1 (86 kDa), Tpz1 (61 kDa), Pot1 (67 kDa), Poz1 (34 kDa), and Rap1 (84 kDa).
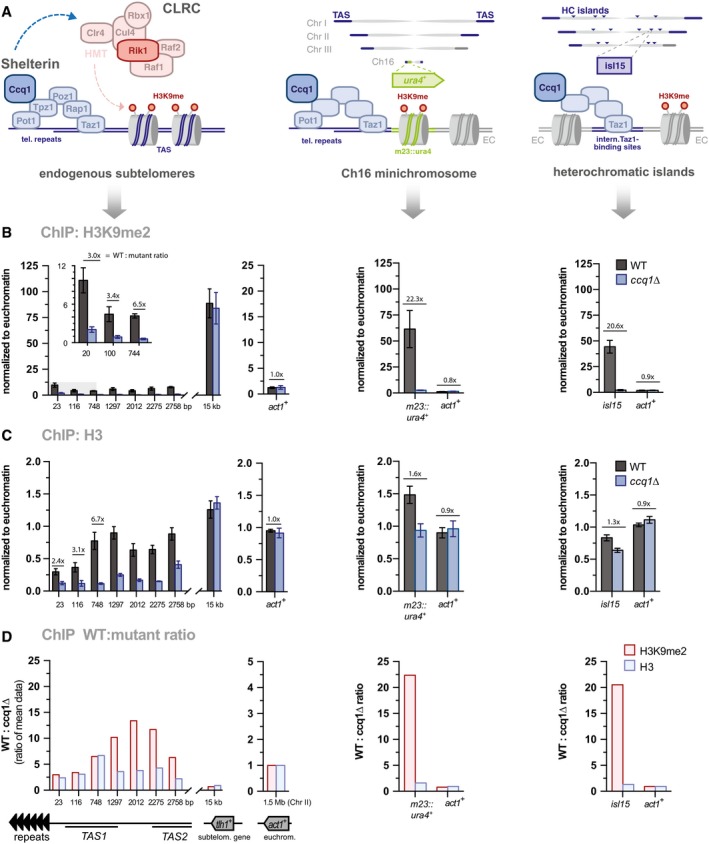
- A
Left: Schemes of CLRC and the shelterin complex. Middle and right: Scheme depicting the position of the ura4 + reporter gene and heterochromatin islands relative to telomeric repeats and TAS regions; note that the minichromosome Ch16 does not contain TAS.
- B, C
ChIP‐qPCR analyses of H3K9me2 (B) and H3 (C) in WT and ccq1Δ cells (n = 3 independent experiments). Data are normalized to input and to the average of three euchromatic loci (EC) as internal control (see Materials and Methods) and represented as mean ± SEM.
- D
Fold change of WT over ccq1Δ for H3K9me2 and H3 levels (see Materials and Methods).
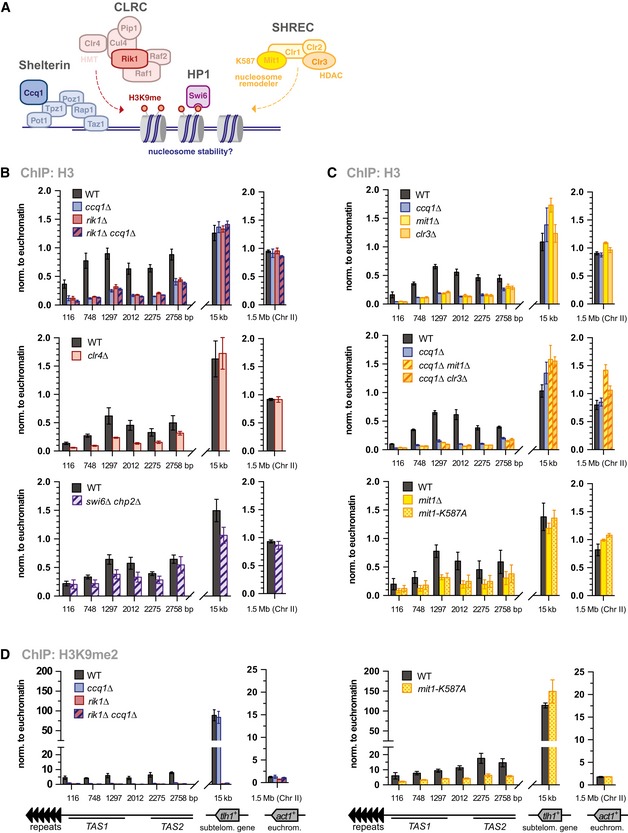
- A
Scheme of shelterin, CLRC and SHREC complexes, and Swi6HP1 bound to H3K9me2.
- B–D
ChIP‐qPCR analyses of H3 and H3K9me2 in WT and mutants as indicated (n = 3 independent experiments). Data are represented as mean ± SEM (normalization as in Fig 2B and C).
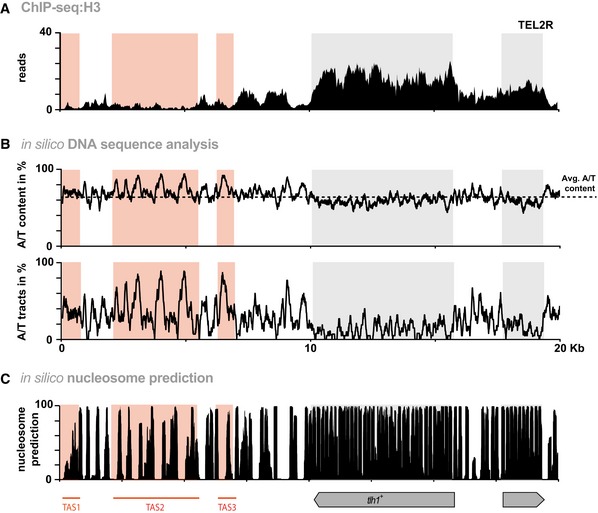
ChIP‐seq reads mapped to TEL2R (shown in reverse orientation for consistency).
In silico analyses of TEL2R DNA sequence. Black line in top panel shows A/T content. Dotted line represents average A/T content in S. pombe (
www.pombase.org ). Bottom panel shows the percentage of poly[dA:dT] tracts (defined as a sequence of 5 or more nucleotides consisting only of A or T). Red and gray shaded areas show TAS regions and subtelomeric genes, respectively.Nucleosome prediction for TEL2R using prediction algorithm 33.
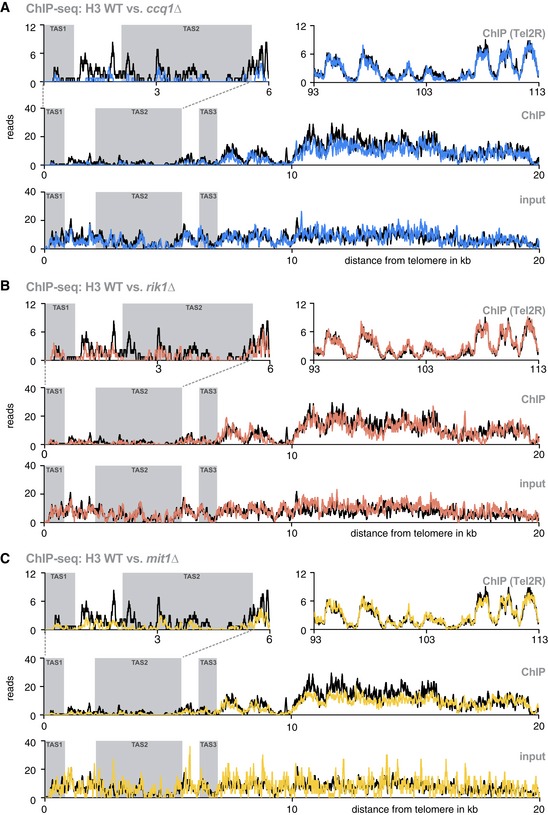
- A–C
ChIP‐seq reads mapped to TEL2R (shown in reverse orientation for consistency) for WT (black) vs. ccq1Δ (blue) (A), rik1Δ (red) (B), and mit1Δ (yellow) cells (C). Shown are reads of ChIP data for TAS1 and TAS2 (top left panel), a nucleosome‐free region reported by Tashiro et al 12 (top right panel), and the subtelomeres including the TAS1‐3 and tlh1 + (middle panel). The bottom panel shows input samples for the same region. Gray shaded areas are TAS regions.
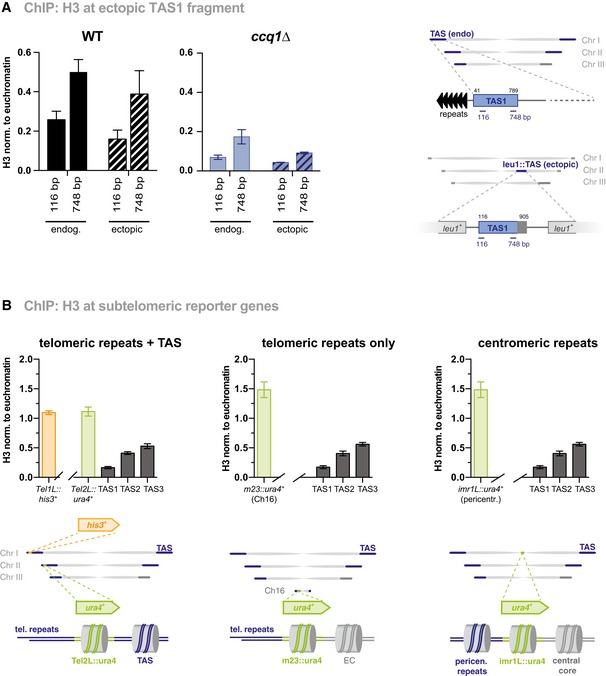
ChIP‐qPCR analysis of H3 at endogenous TAS and a fragment spanning the TAS region from 115 to 905 bp (relative to telomeric repeat) inserted into the leu1 + locus (see scheme). The TAS fragment was inserted into a strain that lacks endogenous TAS (see text) 12. Shown are ChIP analyses for WT (left) and ccq1∆ (right; note different the scale of the y‐axis) (n = 9–10 independent experiments except for ectopic TAS in ccq1∆ strain where n = 3).
ChIP‐qPCR analysis of H3 at reporter genes (ura4 + and his3 +) at various chromosomal locations (see schemes) in WT cells. TAS1, TAS2, and TAS3 correspond to position 116, 2,851, and 6,291 bp (relative to telomeric repeats), respectively (n = 3 independent experiments).
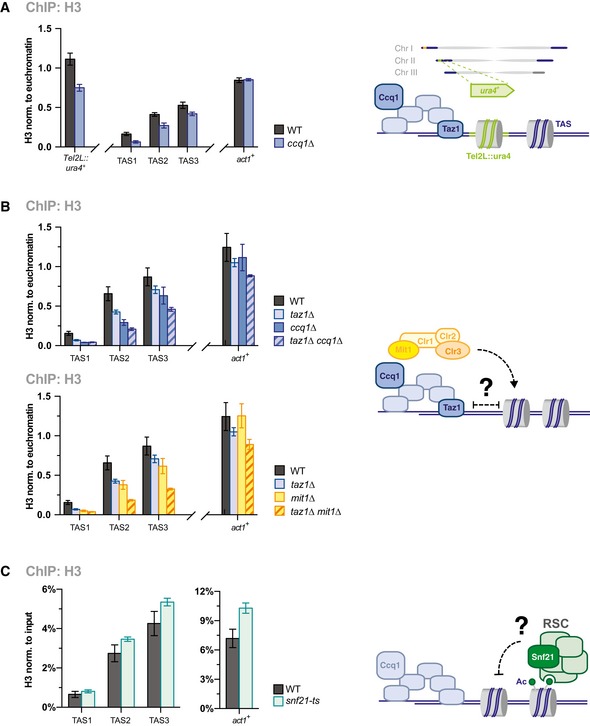
ChIP‐qPCR analysis of H3 at reporter gene (ura4 +) and TAS regions WT and ccq1Δ strains (see scheme) (n = 3 independent experiments).
ChIP‐qPCR analysis of H3 in WT, ccq1Δ, and taz1Δ (top) or mit1Δ (bottom) and corresponding double mutants (see scheme) (n = 3 independent experiments).
ChIP‐qPCR analysis of H3 in WT and snf21‐ts cells (see scheme) (n = 3 independent experiments each derived from 2 to 3 parallel ChIP samples).
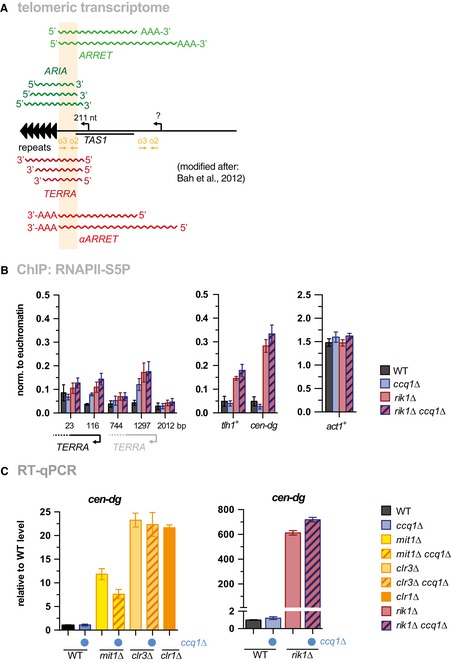
Scheme of telomeric transcripts (modified after 36). Whereas ARRET and αARRET RNAs have a poly‐A tail, only a small percentage of TERRA and ARIA transcripts are poly‐adenylated. The amplicon (primers o2 + o3) used for RT–qPCR and ChIP‐qPCR analysis anneals to all telomeric transcripts without discriminating strand specificity or shorter species that lack transcribed parts from the telomeric repeats. For simplicity, we refer to these transcripts as “TERRA”. An identical sequence of the o2/o3 amplicon is present in a telomere‐distal region, but it is unknown whether this region also contains transcription start sites.
ChIP‐qPCR analysis of RNAPII‐S5P in WT, ccq1Δ, rik1Δ, and the corresponding double mutant (n = 3 independent experiments).
Ccq1 is not involved in silencing of pericentromeric heterochromatin. RT–qPCR analysis of transcript levels of cen‐dg in WT, ccq1Δ, mit1Δ, clr3Δ, rik1Δ, double mutants with ccq1Δ (indicated by blue dot), and clr1Δ cells (n = 3 independent experiments)
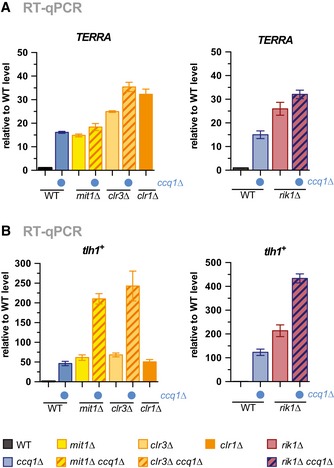
- A, B
RT–qPCR analysis of transcript levels of TERRA (A) and tlh1 + (B) in WT strain and mutants as indicated (double mutants with ccq1Δ are indicated by blue dot). Data are represented as mean ± SEM from three independent experiments and shown relative to WT level (see Materials and Methods).
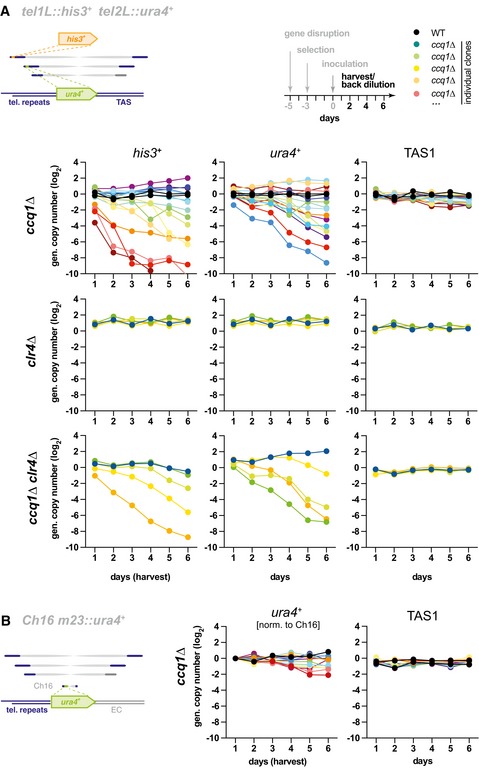
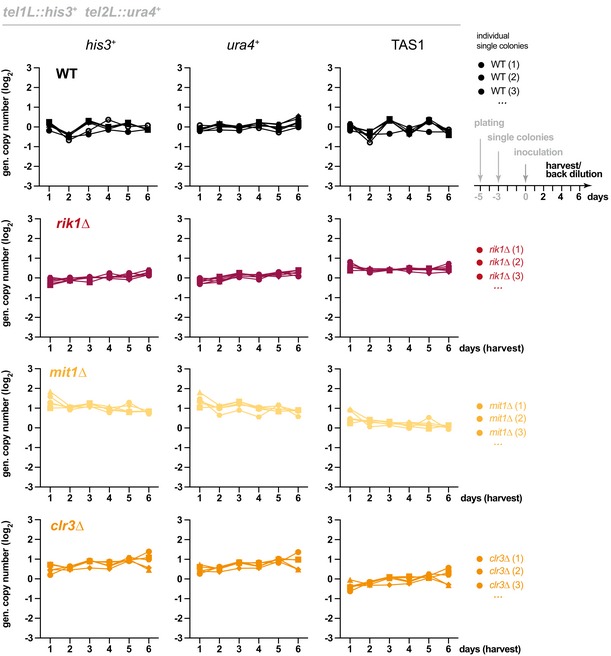
Genomic copy number of his3 +, ura4 +, and TAS1 in indicated strains harboring the reporter genes tel1L::his3 + and tel2L::ura4 +. Cultures from individual WT and freshly generated knockout clones (ccq1Δ, n = 16; clr4Δ, n = 5; ccq1Δ clr4Δ, n = 5) were pre‐grown on selective media for several days and inoculated at day 0 to grow in liquid media with regular back‐dilution (every 24 h, approximately 7 generations). Samples were taken at indicated harvest times, and relative copy numbers of genomic regions were assessed by qPCR (normalization against intrachromosomal loci). Black and rainbow color lines indicate WT strains and individual ccq1 + deletion mutants (clones), respectively.
qPCR analysis as in (A) but with strains harboring the minichromosome Ch16 m23::ura4 + (ccq1Δ, n = 14). Since Ch16 is unstable in ccq1Δ cells (see Appendix Fig S3A and B and 37, m23::ura4 + has been normalized to minichromosome levels (details on normalization, see Materials and Methods). Note that Ch16 does not harbor TAS sequences.
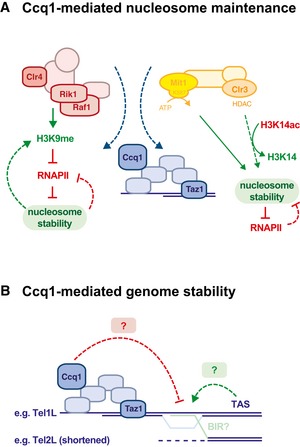
Ccq1 recruits CLRC and SHREC to subtelomeric chromatin, which counteracts nucleosome instability conferred by the DNA sequence of TAS. CLRC may contribute to nucleosome stability indirectly by depositing H3K9me, which prevents access to RNAPII and, thus, nucleosome eviction through transcription. Conversely, stabilized nucleosomes will contribute to maintaining critical levels of H3K9me at TAS. SHREC may contribute to nucleosome stability directly through nucleosome positioning via its ATP‐dependent remodeler activity conferred by Mit1; indirectly through histone deacetylation via the HDAC moiety Clr3, thereby preventing open chromatin structure and access to RNAPII.
TAS or other DNA sequences may represent fragile sites causing DNA breaks through replication fork collapse or the recruitment of unknown factors causing genome instability. Ccq1 may prevent genomic instability by binding to subtelomeric chromatin, thereby stabilizing fragile sites and/or competing with the binding of destabilizing factors. Homologous sequences present in the TAS may promote recombination by BIR (break‐induced replication) or similar mechanisms.
References
-
- Cooper JP, Nimmo ER, Allshire RC, Cech TR (1997) Regulation of telomere length and function by a Myb‐domain protein in fission yeast. Nature 385: 744–747 - PubMed
-
- Miyoshi T, Kanoh J, Saito M, Ishikawa F (2008) Fission yeast Pot1‐Tpp1 protects telomeres and regulates telomere length. Science 320: 1341–1344 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
