

Evolution at two time frames: Polymorphisms from an ancient singular divergence event fuel contemporary parallel evolution
- PMID: 30422983
- PMCID: PMC6258555
- DOI: 10.1371/journal.pgen.1007796
Evolution at two time frames: Polymorphisms from an ancient singular divergence event fuel contemporary parallel evolution
Abstract
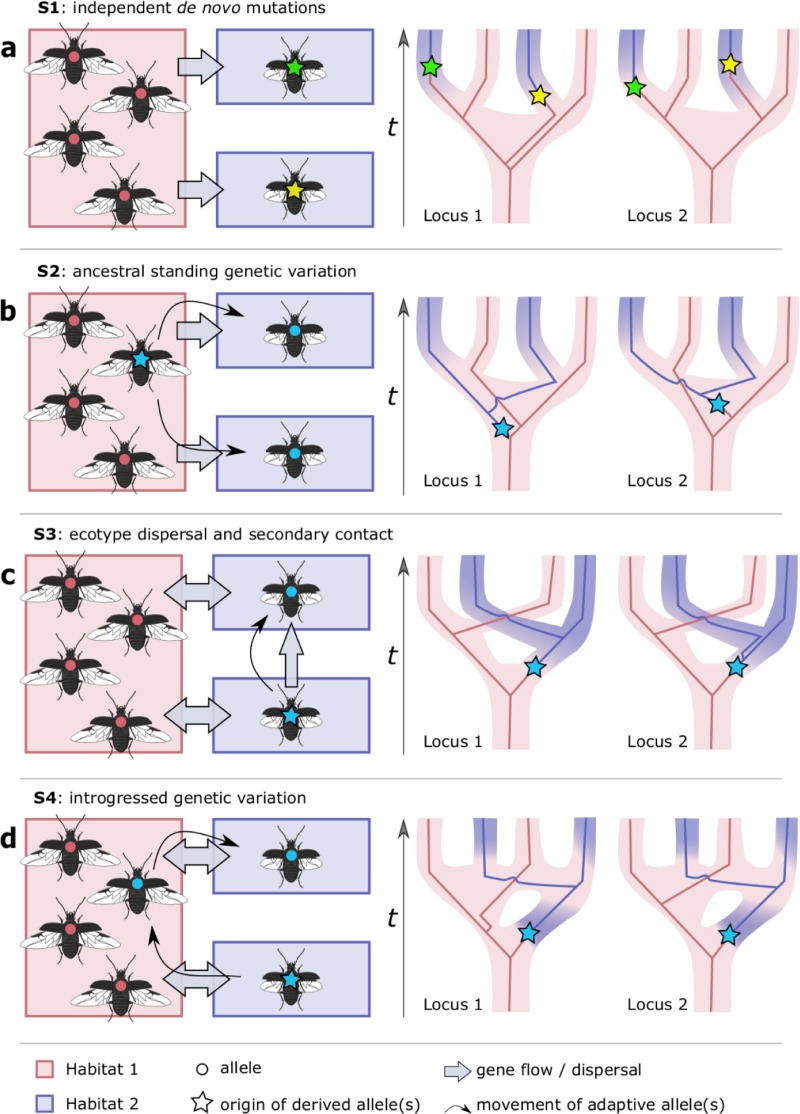
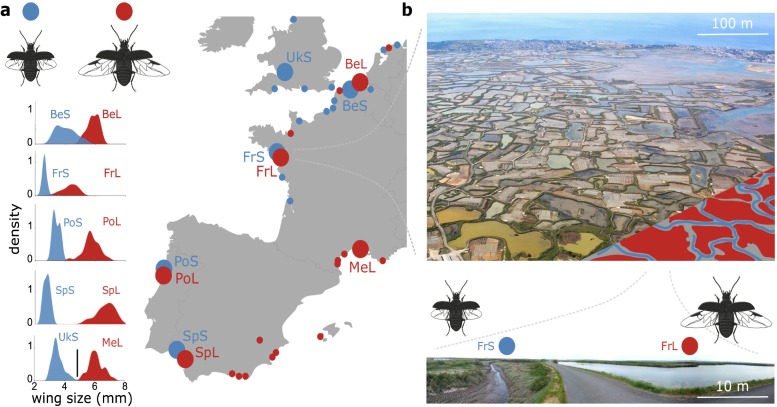
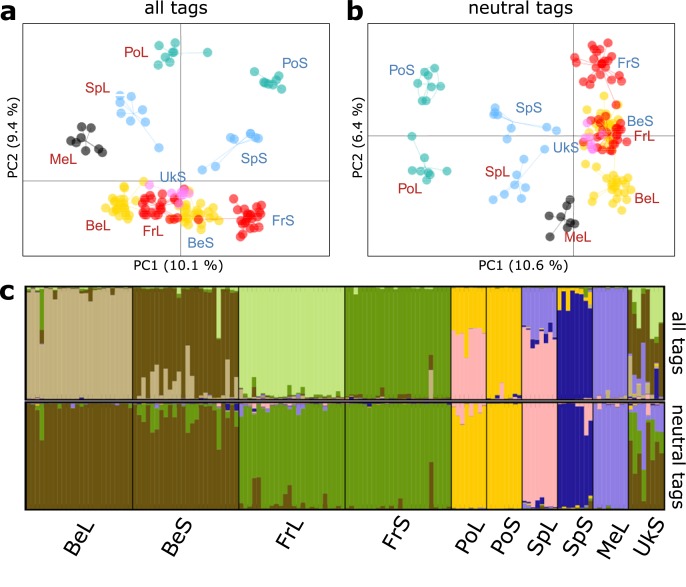
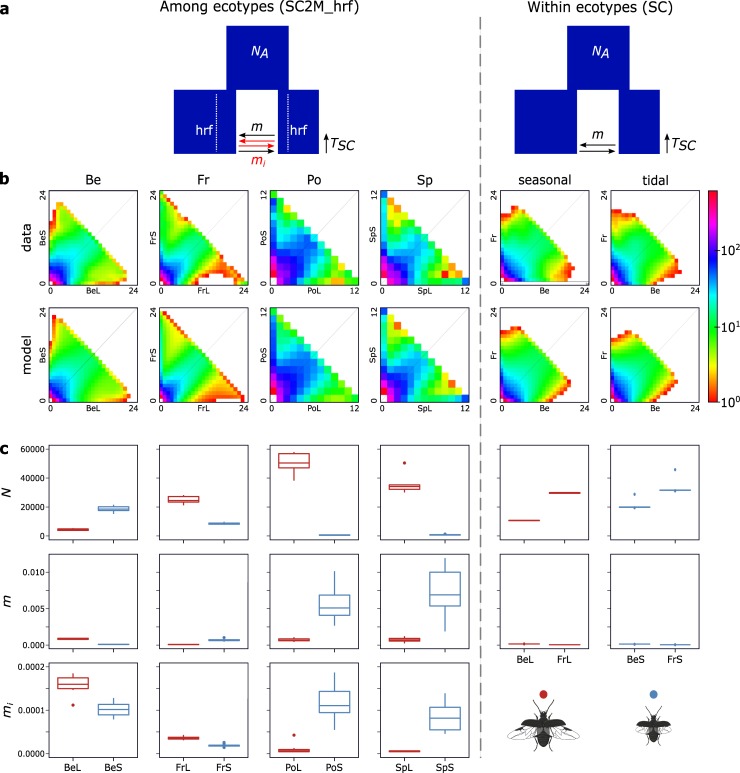
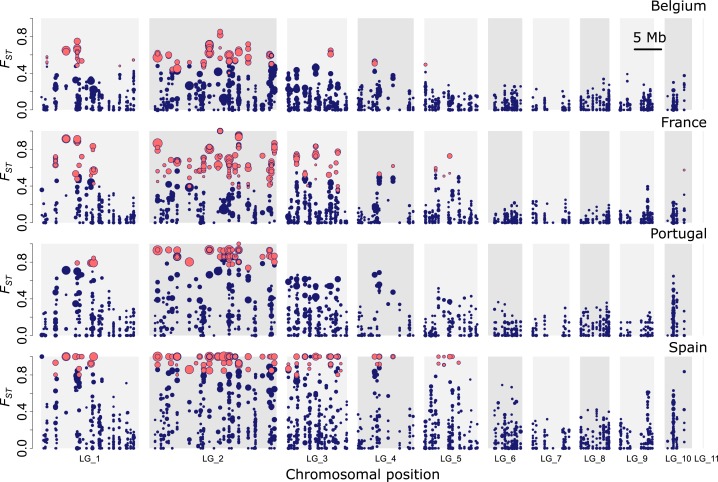
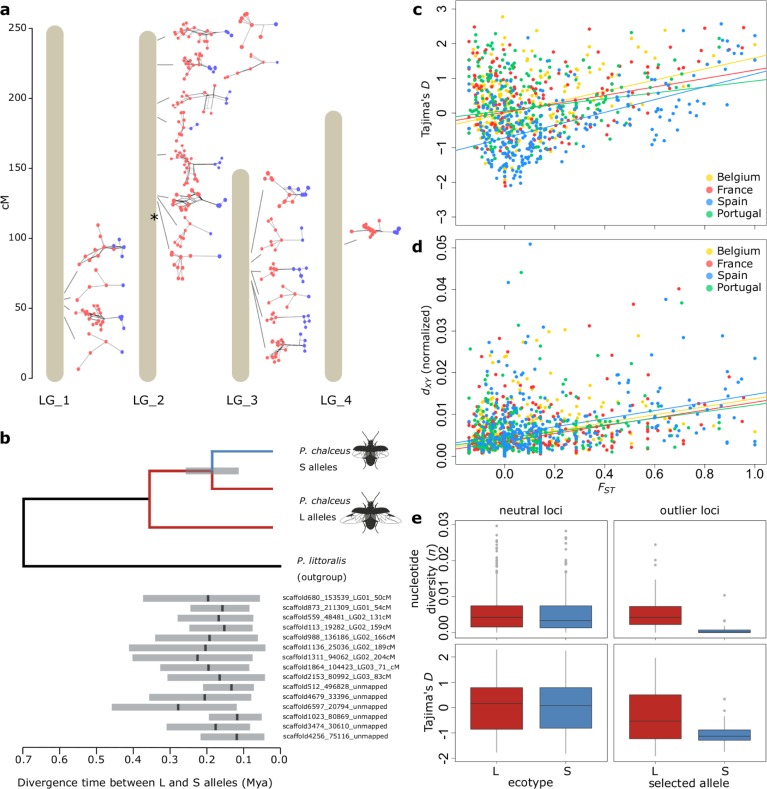
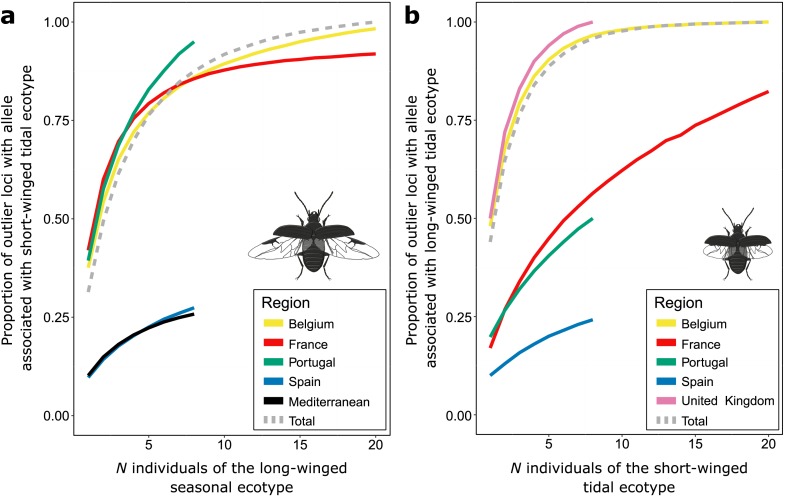
When environments change, populations may adapt surprisingly fast, repeatedly and even at microgeographic scales. There is increasing evidence that such cases of rapid parallel evolution are fueled by standing genetic variation, but the source of this genetic variation remains poorly understood. In the saltmarsh beetle Pogonus chalceus, short-winged 'tidal' and long-winged 'seasonal' ecotypes have diverged in response to contrasting hydrological regimes and can be repeatedly found along the Atlantic European coast. By analyzing genomic variation across the beetles' distribution, we reveal that alleles selected in the tidal ecotype are spread across the genome and evolved during a singular and, likely, geographically isolated divergence event, within the last 190 Kya. Due to subsequent admixture, the ancient and differentially selected alleles are currently polymorphic in most populations across its range, which could potentially allow for the fast evolution of one ecotype from a small number of random individuals, as low as 5 to 15, from a population of the other ecotype. Our results suggest that cases of fast parallel ecological divergence can be the result of evolution at two different time frames: divergence in the past, followed by repeated selection on the same divergently evolved alleles after admixture. These findings highlight the importance of an ancient and, likely, allopatric divergence event for driving the rate and direction of contemporary fast evolution under gene flow. This mechanism is potentially driven by periods of geographic isolation imposed by large-scale environmental changes such as glacial cycles.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Schluter D. Evidence for ecological speciation and its alternative. Science. 2009;323: 737–741. 10.1126/science.1160006 - DOI - PubMed
-
- Nosil P. Ecological Speciation. Oxford, UK: Oxford University Press; 2012.
-
- Coyne JA, Orr HA. Speciation. Sunderland, MA, USA: Sinauer Associates; 2004.
-
- Feder JL, Flaxman SM, Egan SP, Comeault A a., Nosil P. Geographic mode of speciation and genomic divergence. Annu Rev Ecol Evol Syst. 2013;44: 73–97.
-
- Arnegard ME, McGee MD, Matthews B, Marchinko KB, Conte GL, Kabir S, et al. Genetics of ecological divergence during speciation. Nature. Nature Publishing Group; 2014;511: 307–311. 10.1038/nature13301 - DOI - PMC - PubMed
Publication types
MeSH terms
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
