

High-Throughput Stability Screening of Neoantigen/HLA Complexes Improves Immunogenicity Predictions
- PMID: 30425106
- PMCID: PMC6324732
- DOI: 10.1158/2326-6066.CIR-18-0395
High-Throughput Stability Screening of Neoantigen/HLA Complexes Improves Immunogenicity Predictions
Abstract
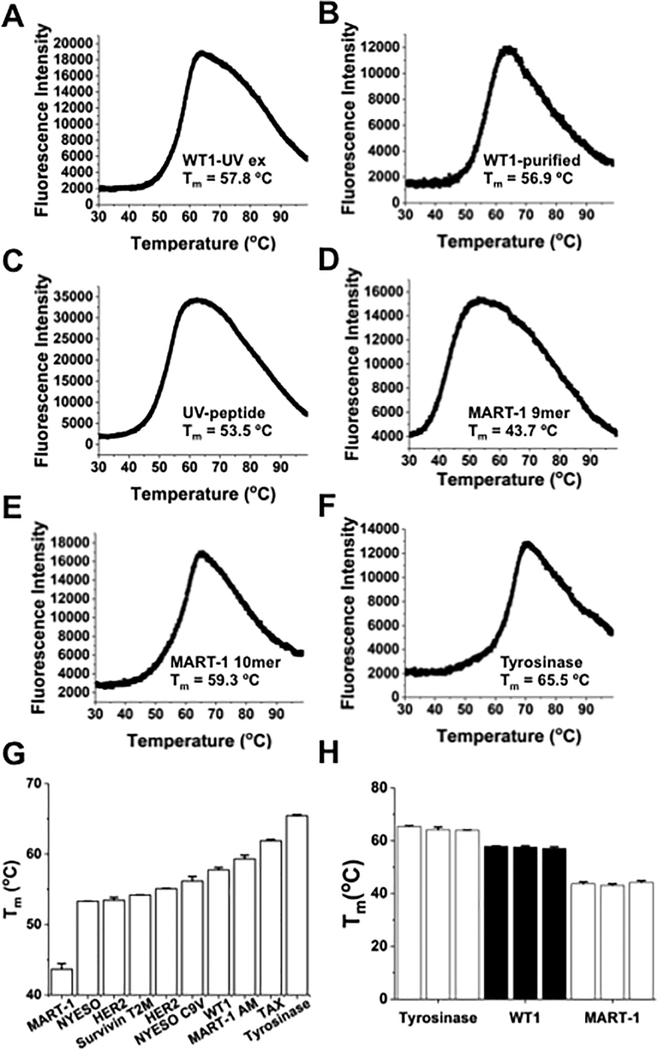
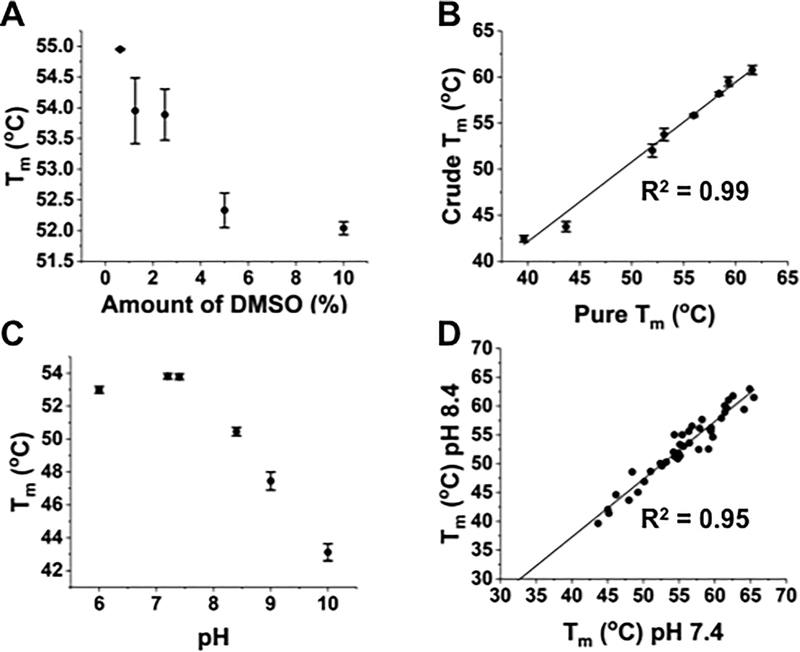
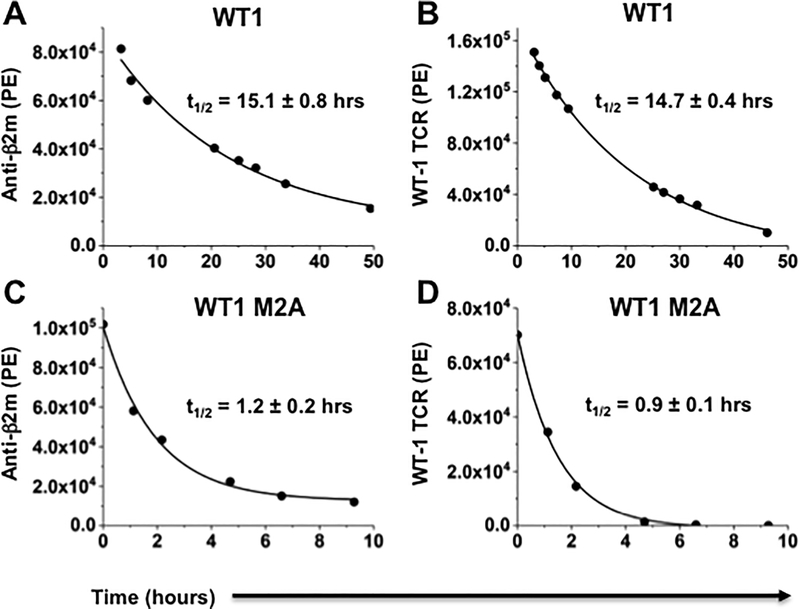
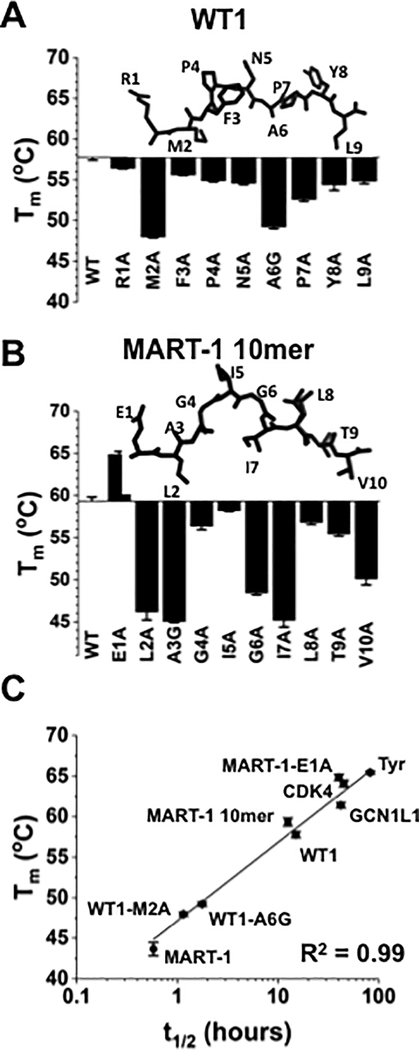
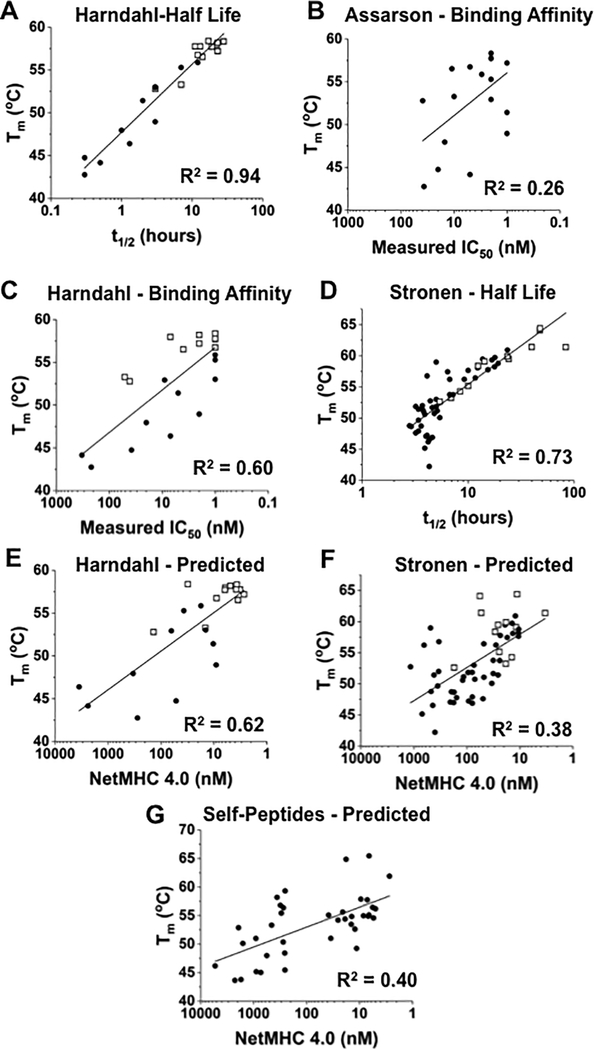
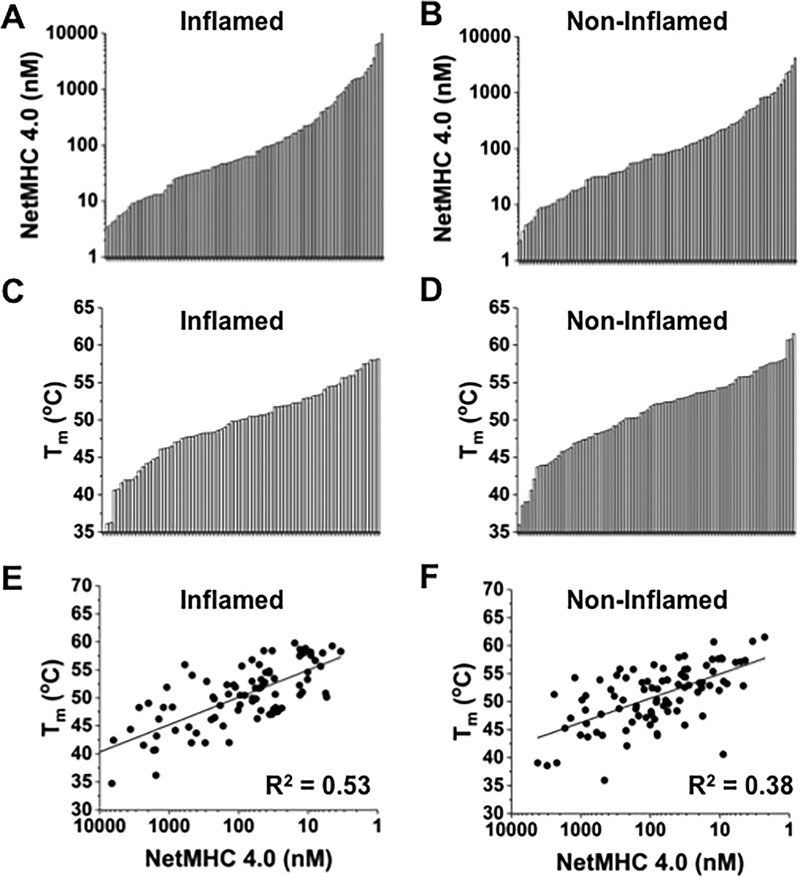
Mutated peptides (neoantigens) from a patient's cancer genome can serve as targets for T-cell immunity, but identifying which peptides can be presented by an MHC molecule and elicit T cells has been difficult. Although algorithms that predict MHC binding exist, they are not yet able to distinguish experimental differences in half-lives of the complexes (an immunologically relevant parameter, referred to here as kinetic stability). Improvement in determining actual neoantigen peptide/MHC stability could be important, as only a small fraction of peptides in most current vaccines are capable of eliciting CD8+ T-cell responses. Here, we used a rapid, high-throughput method to experimentally determine peptide/HLA thermal stability on a scale that will be necessary for analysis of neoantigens from thousands of patients. The method combined the use of UV-cleavable peptide/HLA class I complexes and differential scanning fluorimetry to determine the Tm values of neoantigen complexes. Measured Tm values were accurate and reproducible and were directly proportional to the half-lives of the complexes. Analysis of known HLA-A2-restricted immunogenic peptides showed that Tm values better correlated with immunogenicity than algorithm-predicted binding affinities. We propose that temperature stability information can be used as a guide for the selection of neoantigens in cancer vaccines in order to focus attention on those mutated peptides with the highest probability of being expressed on the cell surface.
©2018 American Association for Cancer Research.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
