

Living at the Frontiers of Life: Extremophiles in Chile and Their Potential for Bioremediation
- PMID: 30425685
- PMCID: PMC6218600
- DOI: 10.3389/fmicb.2018.02309
Living at the Frontiers of Life: Extremophiles in Chile and Their Potential for Bioremediation
Abstract
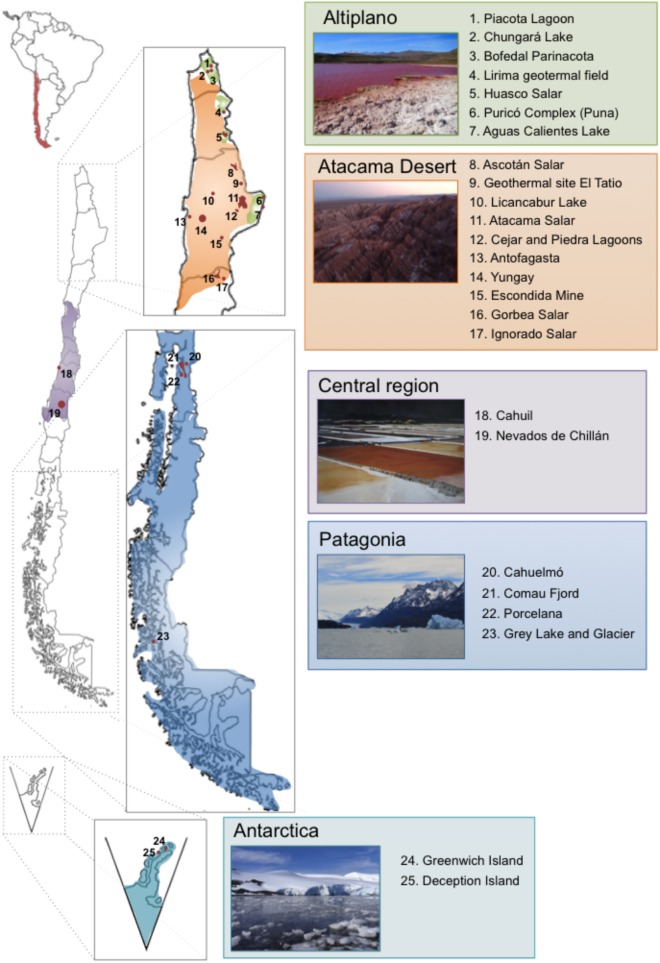
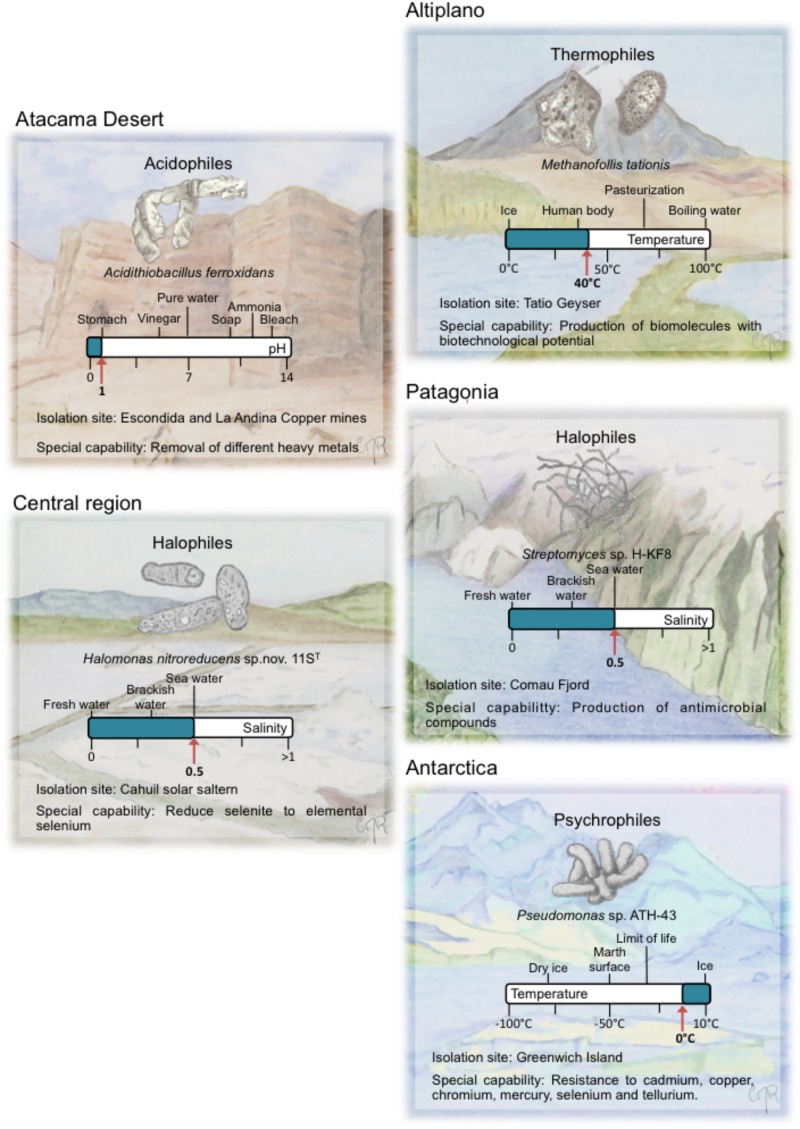
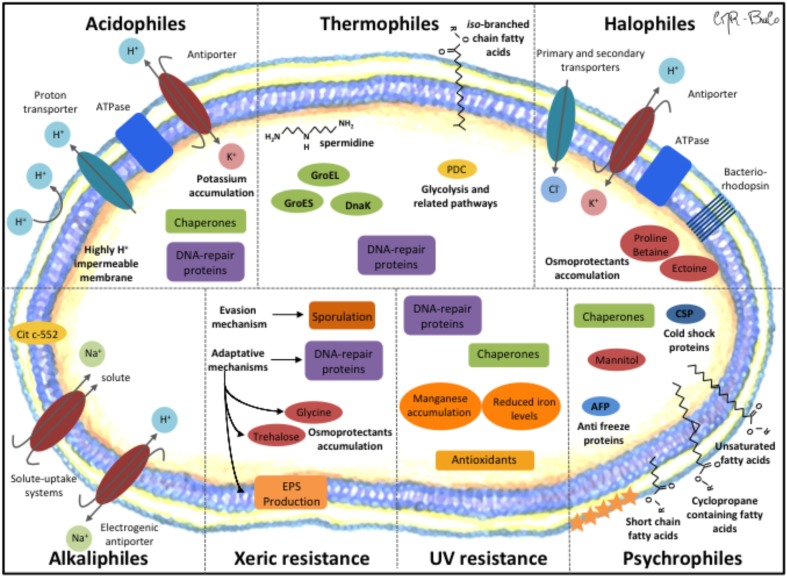
Extremophiles are organisms capable of adjust, survive or thrive in hostile habitats that were previously thought to be adverse or lethal for life. Chile gathers a wide range of extreme environments: salars, geothermal springs, and geysers located at Altiplano and Atacama Desert, salars and cold mountains in Central Chile, and ice fields, cold lakes and fjords, and geothermal sites in Patagonia and Antarctica. The aims of this review are to describe extremophiles that inhabit main extreme biotopes in Chile, and their molecular and physiological capabilities that may be advantageous for bioremediation processes. After briefly describing the main ecological niches of extremophiles along Chilean territory, this review is focused on the microbial diversity and composition of these biotopes microbiomes. Extremophiles have been isolated in diverse zones in Chile that possess extreme conditions such as Altiplano, Atacama Desert, Central Chile, Patagonia, and Antarctica. Interesting extremophiles from Chile with potential biotechnological applications include thermophiles (e.g., Methanofollis tationis from Tatio Geyser), acidophiles (e.g., Acidithiobacillus ferrooxidans, Leptospirillum ferriphilum from Atacama Desert and Central Chile copper ores), halophiles (e.g., Shewanella sp. Asc-3 from Altiplano, Streptomyces sp. HKF-8 from Patagonia), alkaliphiles (Exiguobacterium sp. SH31 from Altiplano), xerotolerant bacteria (S. atacamensis from Atacama Desert), UV- and Gamma-resistant bacteria (Deinococcus peraridilitoris from Atacama Desert) and psychrophiles (e.g., Pseudomonas putida ATH-43 from Antarctica). The molecular and physiological properties of diverse extremophiles from Chile and their application in bioremediation or waste treatments are further discussed. Interestingly, the remarkable adaptative capabilities of extremophiles convert them into an attractive source of catalysts for bioremediation and industrial processes.
Keywords: Altiplano; Antarctica; Atacama Desert; Chile; Patagonia; bioremediation; extremophile.
Figures
References
-
- Abatzopoulos T., Kappas I., Beardmore J., Clegg J. S., Sorgeloos P. (2002). Artemia: Basic and Applied Biology. Dorcrecht: Kluwer Academic Publishers; 10.1007/978-94-017-0791-6 - DOI
-
- Acosta M., Galleguillos P. A., Guajardo M., Demergasso C. (2017). Microbial survey on industrial bioleaching heap by high-throughput 16S sequencing and metagenomics analysis. Solid State Phenom. 266 219–223. 10.4028/www.scientific.net/SSP.262.219 - DOI
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous