

How ERAP1 and ERAP2 Shape the Peptidomes of Disease-Associated MHC-I Proteins
- PMID: 30425713
- PMCID: PMC6219399
- DOI: 10.3389/fimmu.2018.02463
How ERAP1 and ERAP2 Shape the Peptidomes of Disease-Associated MHC-I Proteins
Abstract
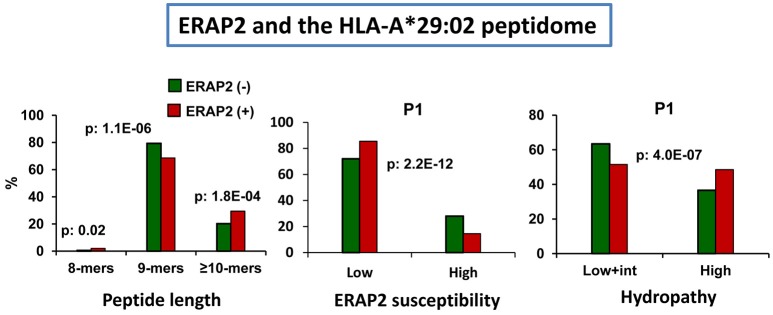
Four inflammatory diseases are strongly associated with Major Histocompatibility Complex class I (MHC-I) molecules: birdshot chorioretinopathy (HLA-A*29:02), ankylosing spondylitis (HLA-B*27), Behçet's disease (HLA-B*51), and psoriasis (HLA-C*06:02). The endoplasmic reticulum aminopeptidases (ERAP) 1 and 2 are also risk factors for these diseases. Since both enzymes are involved in the final processing steps of MHC-I ligands it is reasonable to assume that MHC-I-bound peptides play a significant pathogenetic role. This review will mainly focus on recent studies concerning the effects of ERAP1 and ERAP2 polymorphism and expression on shaping the peptidome of disease-associated MHC-I molecules in live cells. These studies will be discussed in the context of the distinct mechanisms and substrate preferences of both enzymes, their different patterns of genetic association with various diseases, the role of polymorphisms determining changes in enzymatic activity or expression levels, and the distinct peptidomes of disease-associated MHC-I allotypes. ERAP1 and ERAP2 polymorphism and expression induce significant changes in multiple MHC-I-bound peptidomes. These changes are MHC allotype-specific and, without excluding a degree of functional inter-dependence between both enzymes, reflect largely separate roles in their processing of MHC-I ligands. The studies reviewed here provide a molecular basis for the distinct patterns of genetic association of ERAP1 and ERAP2 with disease and for the pathogenetic role of peptides. The allotype-dependent alterations induced on distinct peptidomes may explain that the joint association of both enzymes and unrelated MHC-I alleles influence different pathological outcomes.
Keywords: Behçet's disease; ERAP; MHC; ankylosing spondylitis; antigen processing; inflammatory diseases; psoriasis; uveitis.
Figures

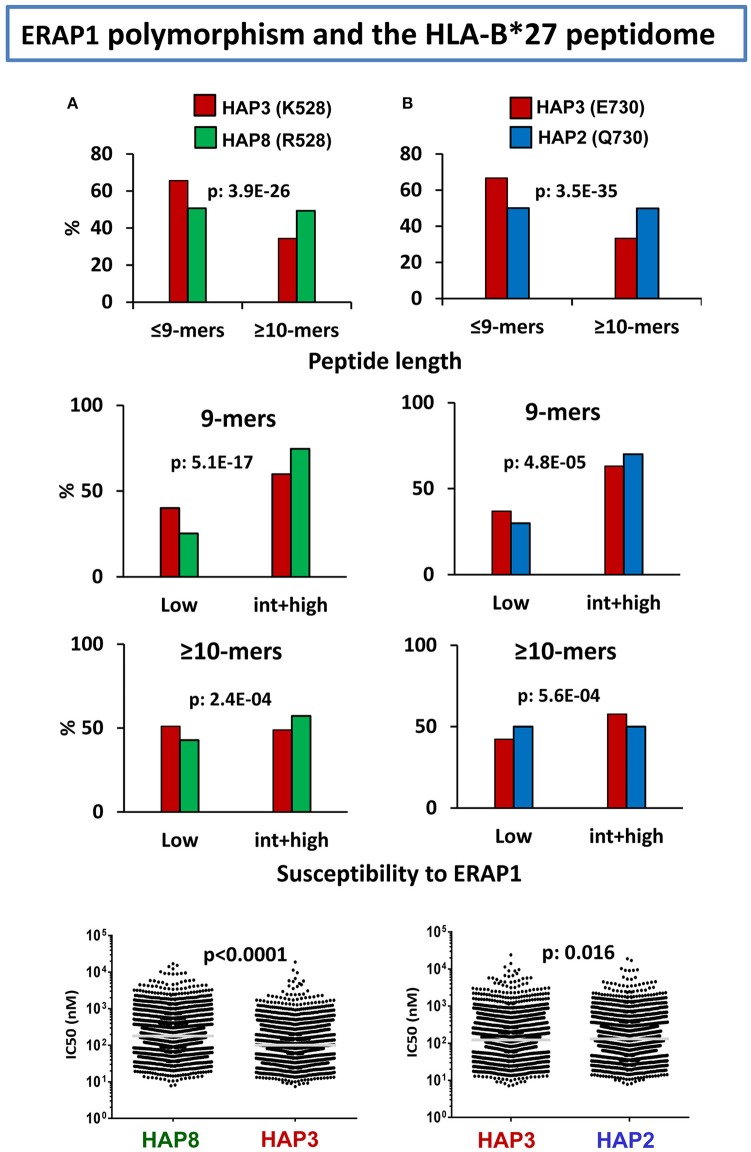
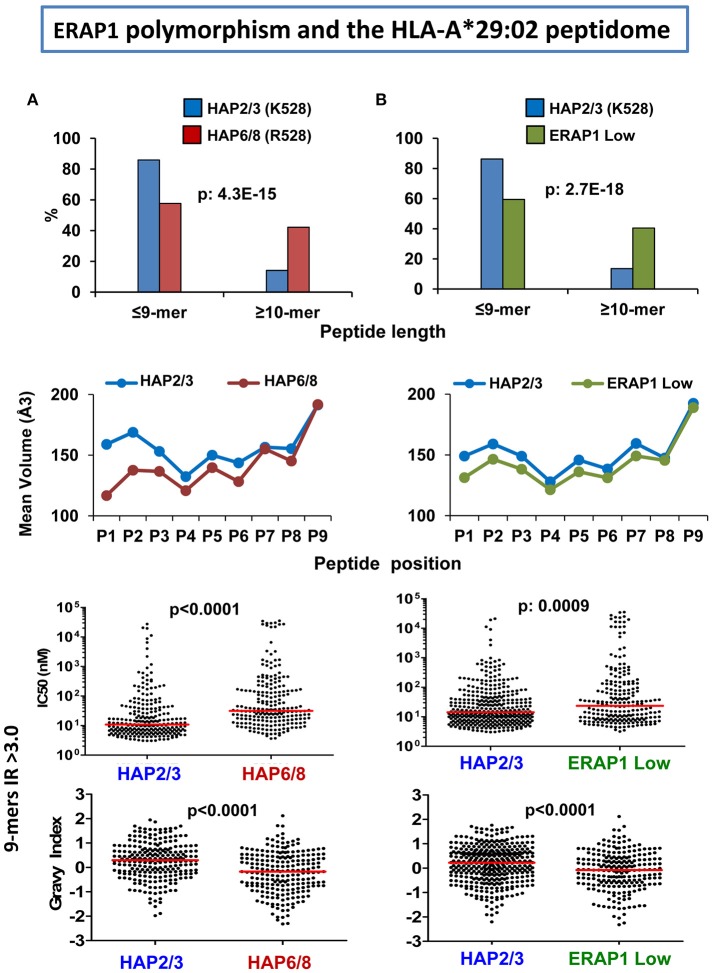
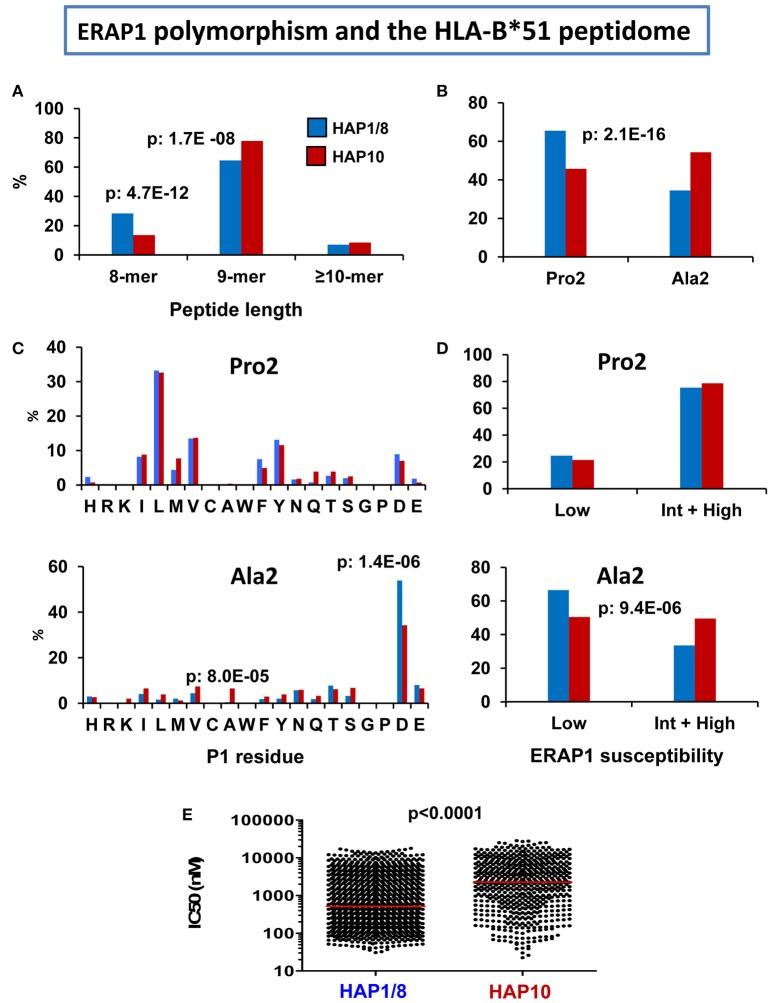
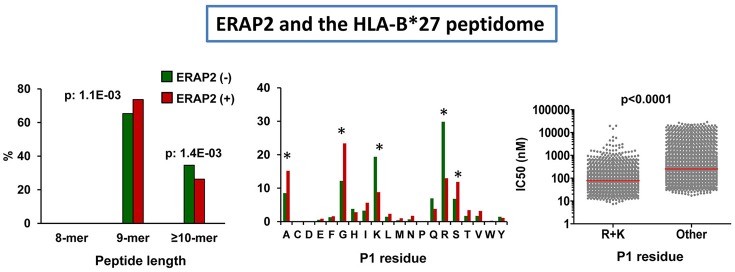
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous