

Analysis of Two New Arabinosyltransferases Belonging to the Carbohydrate-Active Enzyme (CAZY) Glycosyl Transferase Family1 Provides Insights into Disease Resistance and Sugar Donor Specificity
- PMID: 30429223
- PMCID: PMC6354260
- DOI: 10.1105/tpc.18.00641
Analysis of Two New Arabinosyltransferases Belonging to the Carbohydrate-Active Enzyme (CAZY) Glycosyl Transferase Family1 Provides Insights into Disease Resistance and Sugar Donor Specificity
Abstract
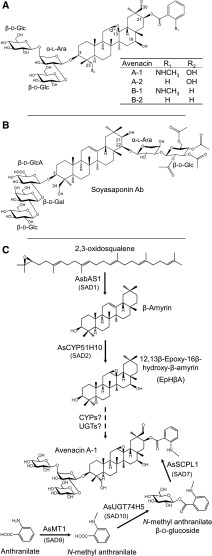
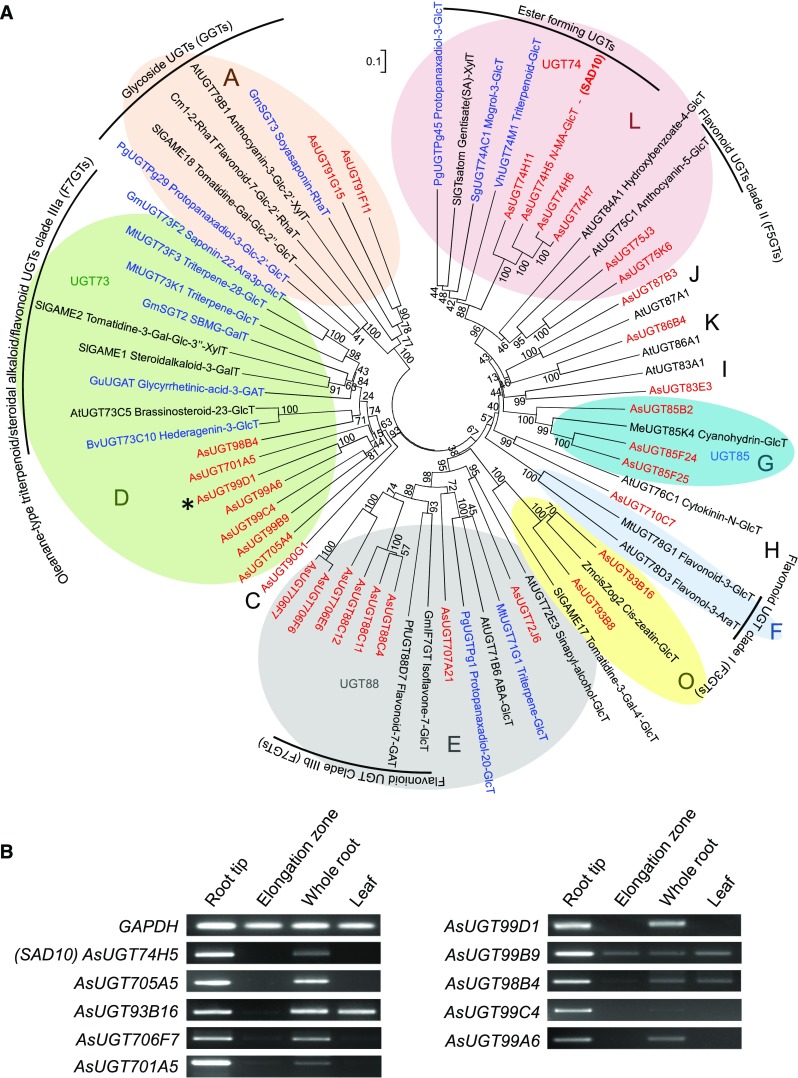
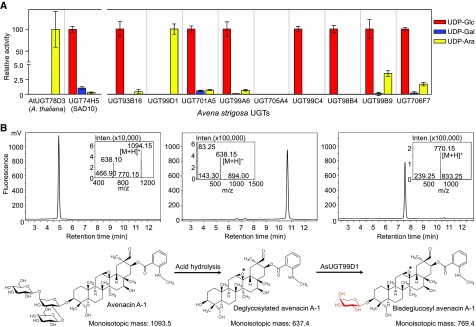
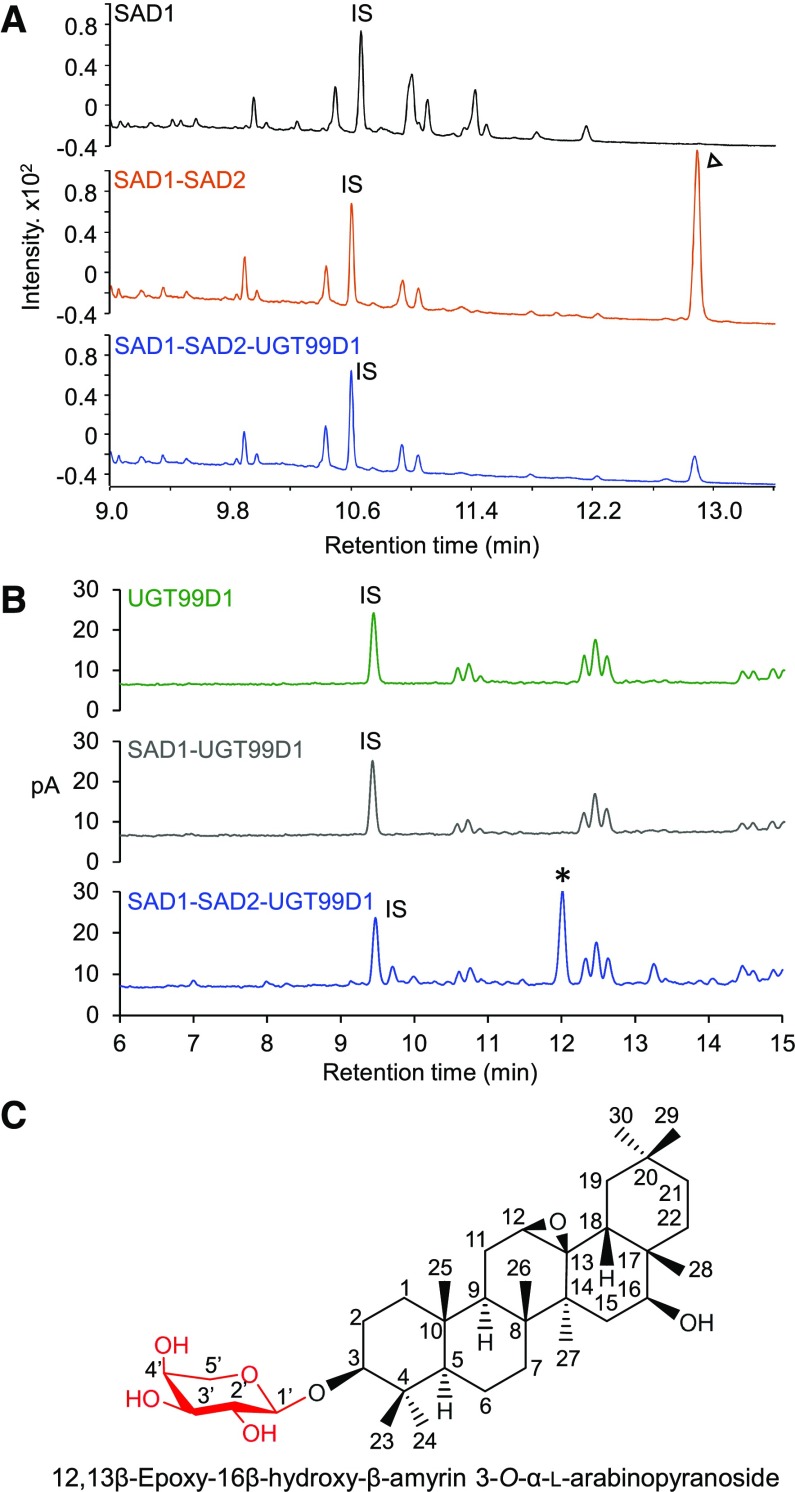
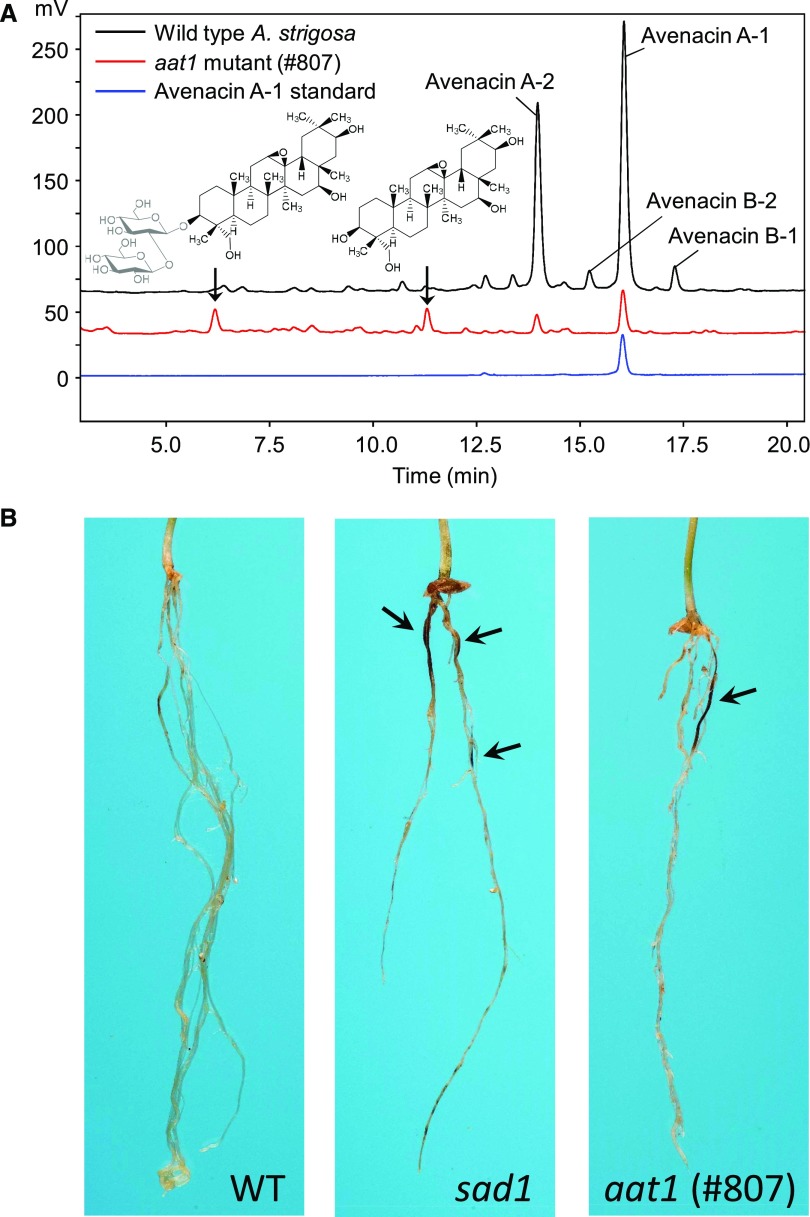
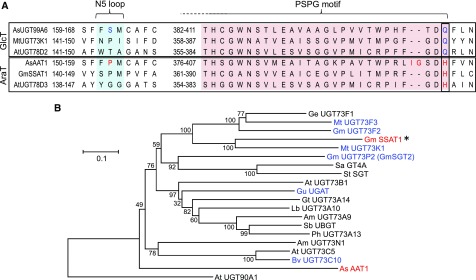
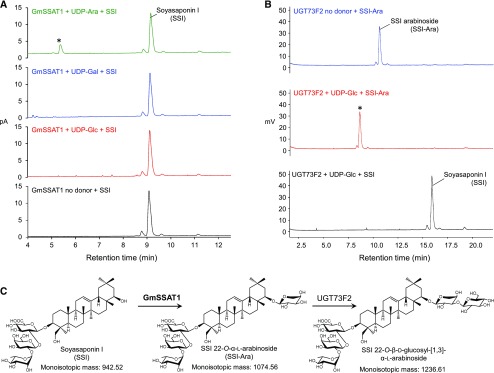
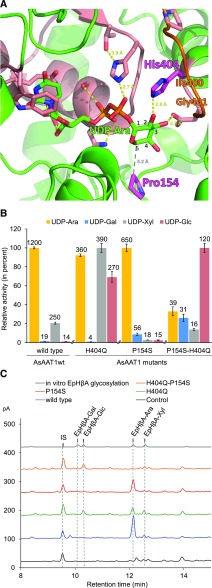
Glycosylation of small molecules is critical for numerous biological processes in plants, including hormone homeostasis, neutralization of xenobiotics, and synthesis and storage of specialized metabolites. Glycosylation of plant natural products is usually performed by uridine diphosphate-dependent glycosyltransferases (UGTs). Triterpene glycosides (saponins) are a large family of plant natural products that determine important agronomic traits such as disease resistance and flavor and have numerous pharmaceutical applications. Most characterized plant natural product UGTs are glucosyltransferases, and little is known about enzymes that add other sugars. Here we report the discovery and characterization of AsAAT1 (UGT99D1), which is required for biosynthesis of the antifungal saponin avenacin A-1 in oat (Avena strigosa). This enzyme adds l-Ara to the triterpene scaffold at the C-3 position, a modification critical for disease resistance. The only previously reported plant natural product arabinosyltransferase is a flavonoid arabinosyltransferase from Arabidopsis (Arabidopsis thaliana). We show that AsAAT1 has high specificity for UDP-β-l-arabinopyranose, identify two amino acids required for sugar donor specificity, and through targeted mutagenesis convert AsAAT1 into a glucosyltransferase. We further identify a second arabinosyltransferase potentially implicated in the biosynthesis of saponins that determine bitterness in soybean (Glycine max). Our investigations suggest independent evolution of UDP-Ara sugar donor specificity in arabinosyltransferases in monocots and eudicots.
© 2018 American Society of Plant Biologists. All rights reserved.
Figures

Similar articles
-
A noncanonical vacuolar sugar transferase required for biosynthesis of antimicrobial defense compounds in oat.Proc Natl Acad Sci U S A. 2019 Dec 26;116(52):27105-27114. doi: 10.1073/pnas.1914652116. Epub 2019 Dec 5. Proc Natl Acad Sci U S A. 2019. PMID: 31806756 Free PMC article.
-
Engineering flavonoid glycosyltransferases for enhanced catalytic efficiency and extended sugar-donor selectivity.Planta. 2013 Oct;238(4):683-93. doi: 10.1007/s00425-013-1922-0. Epub 2013 Jun 26. Planta. 2013. PMID: 23801300
-
Triterpenoid-biosynthetic UDP-glycosyltransferases from plants.Biotechnol Adv. 2019 Nov 15;37(7):107394. doi: 10.1016/j.biotechadv.2019.04.016. Epub 2019 May 9. Biotechnol Adv. 2019. PMID: 31078628 Review.
-
Reconstruction of the Evolutionary Histories of UGT Gene Superfamily in Legumes Clarifies the Functional Divergence of Duplicates in Specialized Metabolism.Int J Mol Sci. 2020 Mar 8;21(5):1855. doi: 10.3390/ijms21051855. Int J Mol Sci. 2020. PMID: 32182686 Free PMC article.
-
The Sweet Side of Plant-Specialized Metabolism.Cold Spring Harb Perspect Biol. 2019 Dec 2;11(12):a034744. doi: 10.1101/cshperspect.a034744. Cold Spring Harb Perspect Biol. 2019. PMID: 31235546 Free PMC article. Review.
Cited by
-
Chromosome-scale genome assembly of Glycyrrhiza uralensis revealed metabolic gene cluster centred specialized metabolites biosynthesis.DNA Res. 2022 Dec 1;29(6):dsac043. doi: 10.1093/dnares/dsac043. DNA Res. 2022. PMID: 36535891 Free PMC article.
-
Advances in steroidal saponins biosynthesis.Planta. 2021 Oct 6;254(5):91. doi: 10.1007/s00425-021-03732-y. Planta. 2021. PMID: 34617240 Review.
-
Glycosyltransferases: Mining, engineering and applications in biosynthesis of glycosylated plant natural products.Synth Syst Biotechnol. 2022 Feb 2;7(1):602-620. doi: 10.1016/j.synbio.2022.01.001. eCollection 2022 Mar. Synth Syst Biotechnol. 2022. PMID: 35261926 Free PMC article.
-
Metabolic pathway genes for editing to enhance multiple disease resistance in plants.J Plant Res. 2022 Nov;135(6):705-722. doi: 10.1007/s10265-022-01409-5. Epub 2022 Aug 29. J Plant Res. 2022. PMID: 36036859
-
Plant terpenoid metabolism co-opts a component of the cell wall biosynthesis machinery.Nat Chem Biol. 2020 Jul;16(7):740-748. doi: 10.1038/s41589-020-0541-x. Epub 2020 May 18. Nat Chem Biol. 2020. PMID: 32424305
References
-
- Achnine L., Huhman D.V., Farag M.A., Sumner L.W., Blount J.W., Dixon R.A. (2005). Genomics-based selection and functional characterization of triterpene glycosyltransferases from the model legume Medicago truncatula. Plant J. 41: 875–887. - PubMed
-
- Augustin J.M., Kuzina V., Andersen S.B., Bak S. (2011). Molecular activities, biosynthesis and evolution of triterpenoid saponins. Phytochemistry 72: 435–457. - PubMed
-
- Augustin J.M., Drok S., Shinoda T., Sanmiya K., Nielsen J.K., Khakimov B., Olsen C.E., Hansen E.H., Kuzina V., Ekstrøm C.T., Hauser T., Bak S. (2012). UDP-glycosyltransferases from the UGT73C subfamily in Barbarea vulgaris catalyze sapogenin 3-O-glucosylation in saponin-mediated insect resistance. Plant Physiol. 160: 1881–1895. - PMC - PubMed
-
- Benkert P., Tosatto S.C., Schomburg D. (2008). QMEAN: A comprehensive scoring function for model quality assessment. Proteins 71: 261–277. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- BBS/E/J/000PR9790/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/J/000C0618/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/J/00000614/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/K005952/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/P012523/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
