

The mitochondrial calcium uniporter underlies metabolic fuel preference in skeletal muscle
- PMID: 30429366
- PMCID: PMC6302934
- DOI: 10.1172/jci.insight.121689
The mitochondrial calcium uniporter underlies metabolic fuel preference in skeletal muscle
Abstract
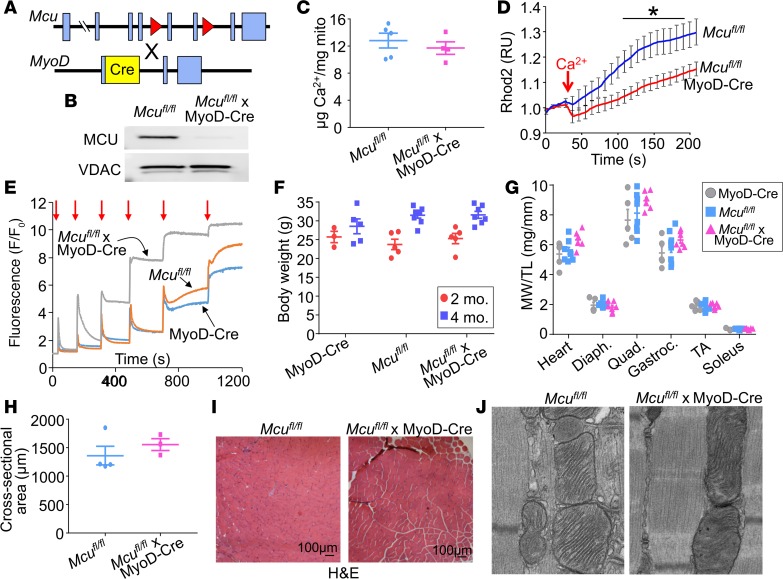
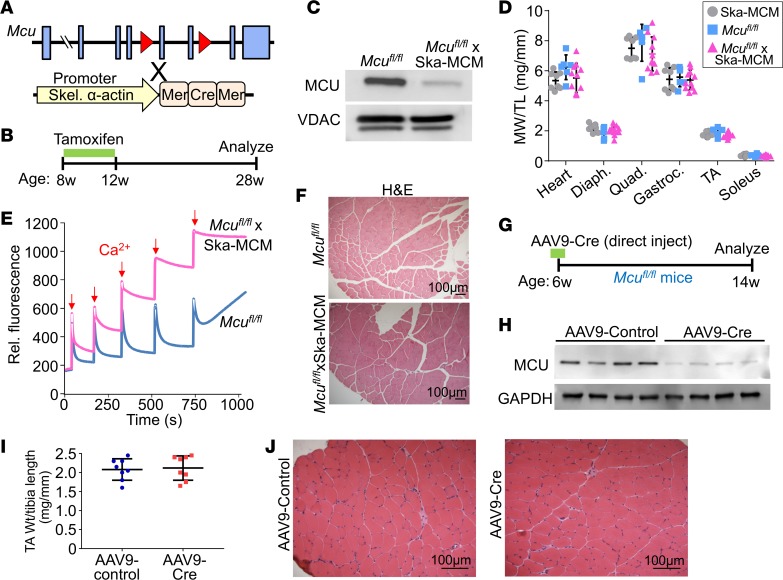
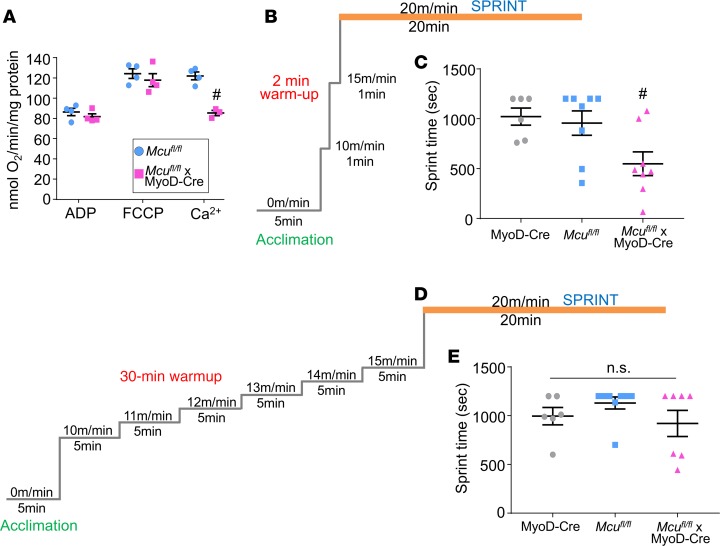
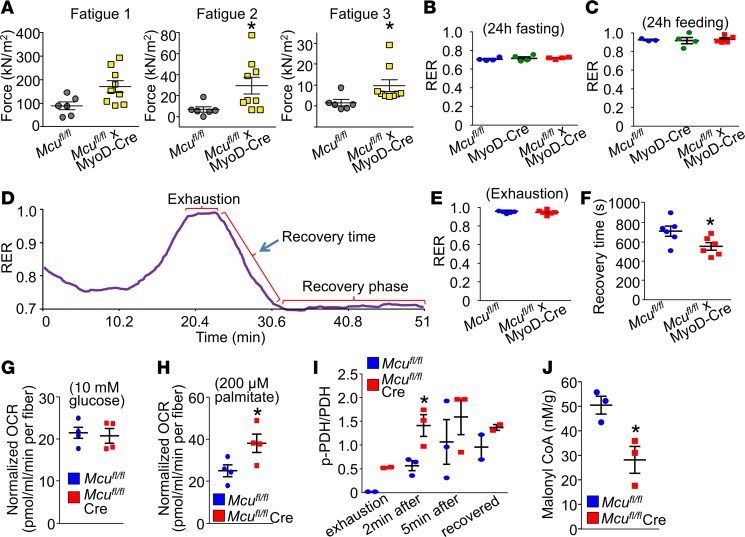
The mitochondrial Ca2+ uniporter (MCU) complex mediates acute mitochondrial Ca2+ influx. In skeletal muscle, MCU links Ca2+ signaling to energy production by directly enhancing the activity of key metabolic enzymes in the mitochondria. Here, we examined the role of MCU in skeletal muscle development and metabolic function by generating mouse models for the targeted deletion of Mcu in embryonic, postnatal, and adult skeletal muscle. Loss of Mcu did not affect muscle growth and maturation or otherwise cause pathology. Skeletal muscle-specific deletion of Mcu in mice also did not affect myofiber intracellular Ca2+ handling, but it did inhibit acute mitochondrial Ca2+ influx and mitochondrial respiration stimulated by Ca2+, resulting in reduced acute exercise performance in mice. However, loss of Mcu also resulted in enhanced muscle performance under conditions of fatigue, with a preferential shift toward fatty acid metabolism, resulting in reduced body fat with aging. Together, these results demonstrate that MCU-mediated mitochondrial Ca2+ regulation underlies skeletal muscle fuel selection at baseline and under enhanced physiological demands, which affects total homeostatic metabolism.
Keywords: Calcium; Cardiology; Mitochondria; Muscle Biology.
Conflict of interest statement
Figures
References
-
- Denton RM. Regulation of mitochondrial dehydrogenases by calcium ions. Biochim Biophys Acta. 2009;1787(11):1309–1316. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
