

Presenilin-mediated cleavage of APP regulates synaptotagmin-7 and presynaptic plasticity
- PMID: 30429473
- PMCID: PMC6235831
- DOI: 10.1038/s41467-018-06813-x
Presenilin-mediated cleavage of APP regulates synaptotagmin-7 and presynaptic plasticity
Abstract
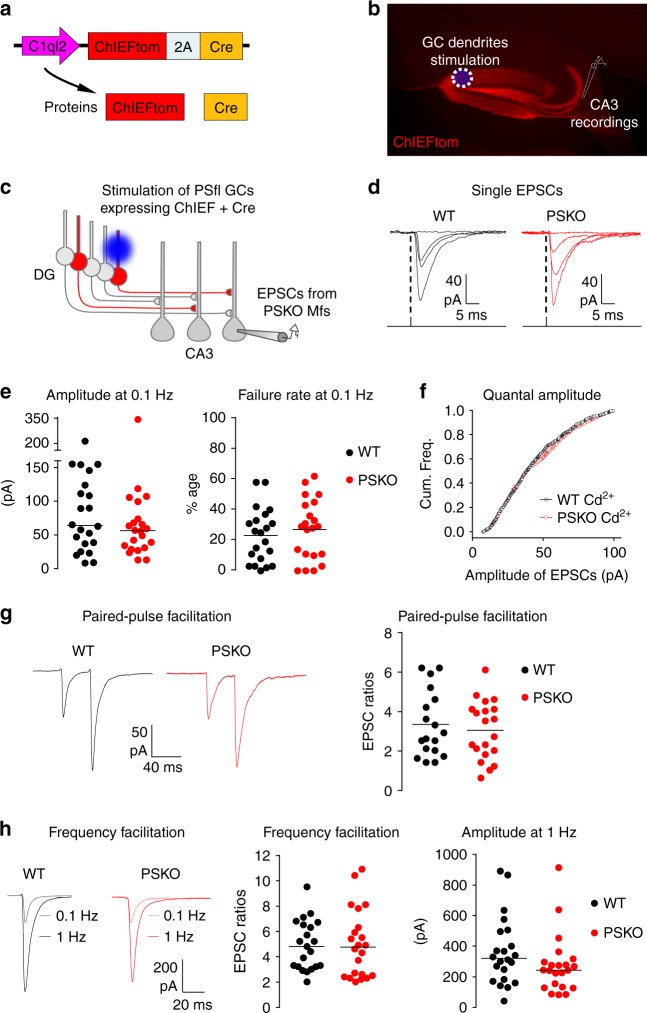
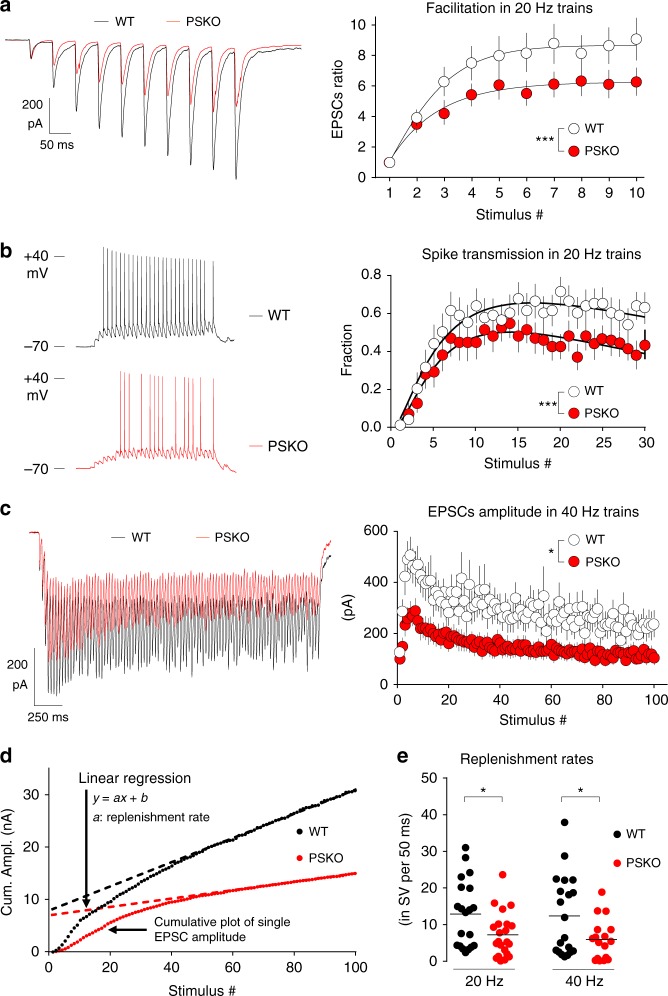
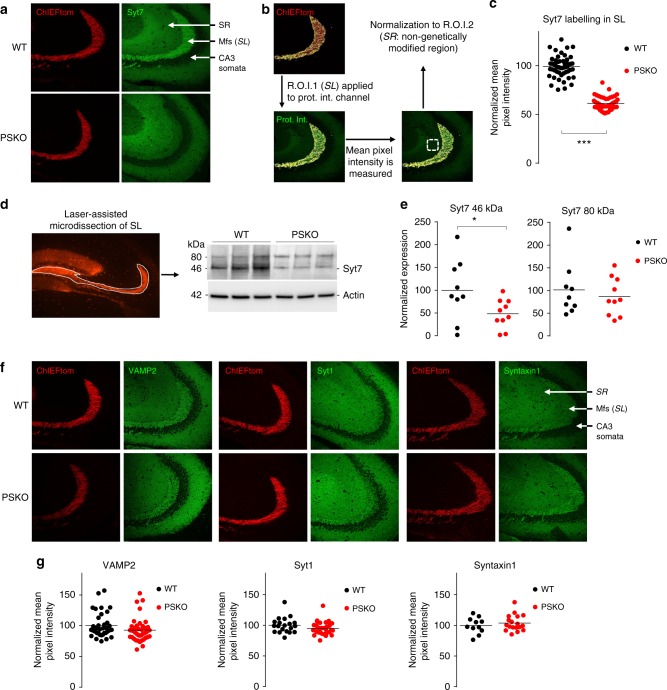
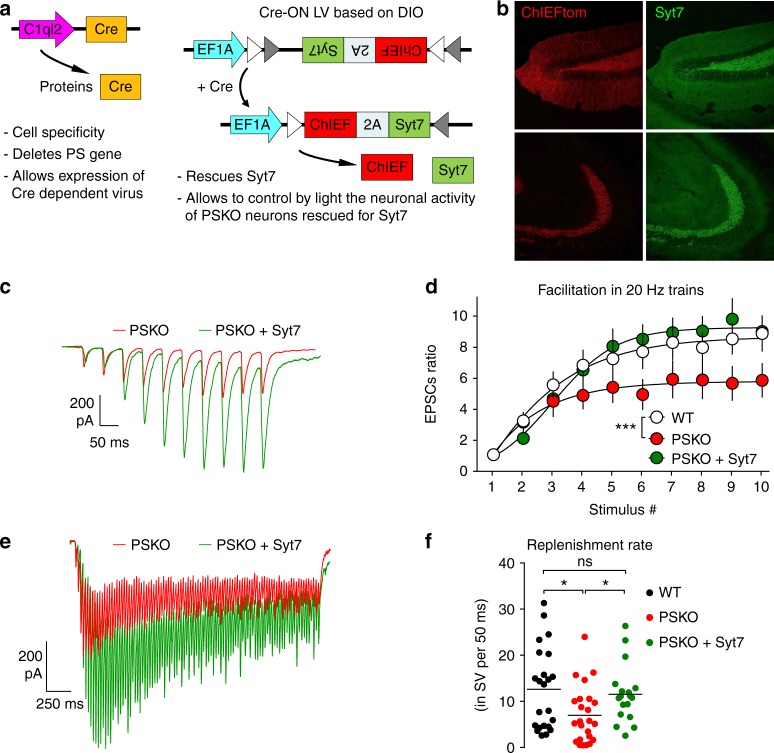
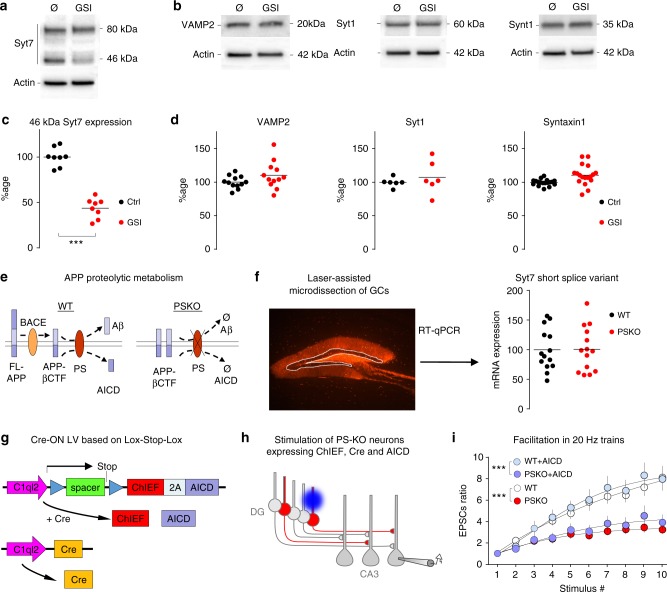
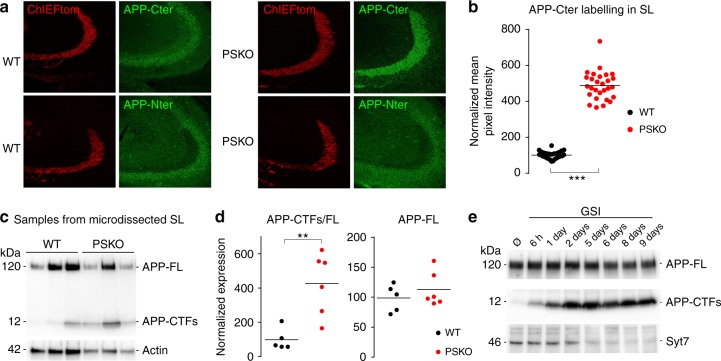
Mutations of the intramembrane protease presenilin (PS) or of its main substrate, the amyloid precursor protein (APP), cause early-onset form of Alzheimer disease. PS and APP interact with proteins of the neurotransmitter release machinery without identified functional consequences. Here we report that genetic deletion of PS markedly decreases the presynaptic levels of the Ca2+ sensor synaptotagmin-7 (Syt7) leading to impaired synaptic facilitation and replenishment of synaptic vesicles. The regulation of Syt7 expression by PS occurs post-transcriptionally and depends on γ-secretase proteolytic activity. It requires the substrate APP as revealed by the combined genetic invalidation of APP and PS1, and in particular the APP-Cterminal fragments which interact with Syt7 and accumulate in synaptic terminals under pharmacological or genetic inhibition of γ-secretase. Thus, we uncover a role of PS in presynaptic mechanisms, through APP cleavage and regulation of Syt7, that highlights aberrant synaptic vesicle processing as a possible new pathway in AD.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
