

Revised role for Hfq bacterial regulator on DNA topology
- PMID: 30429520
- PMCID: PMC6235962
- DOI: 10.1038/s41598-018-35060-9
Revised role for Hfq bacterial regulator on DNA topology
Abstract
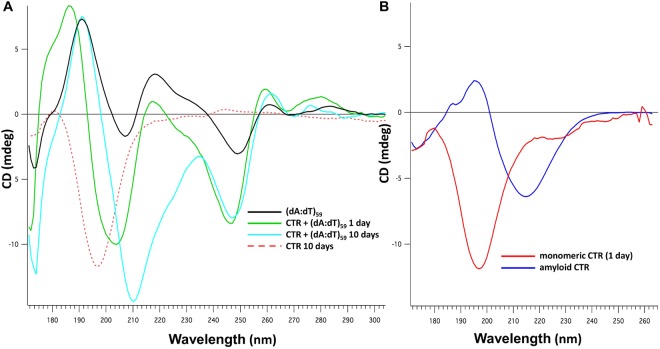
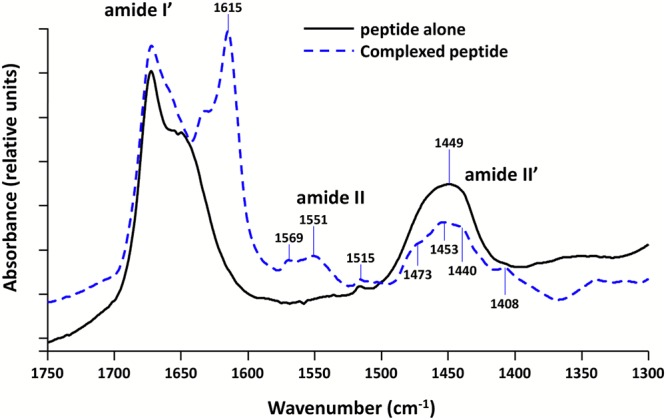
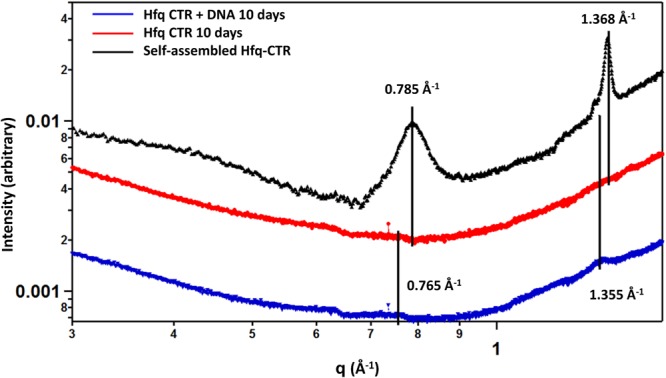
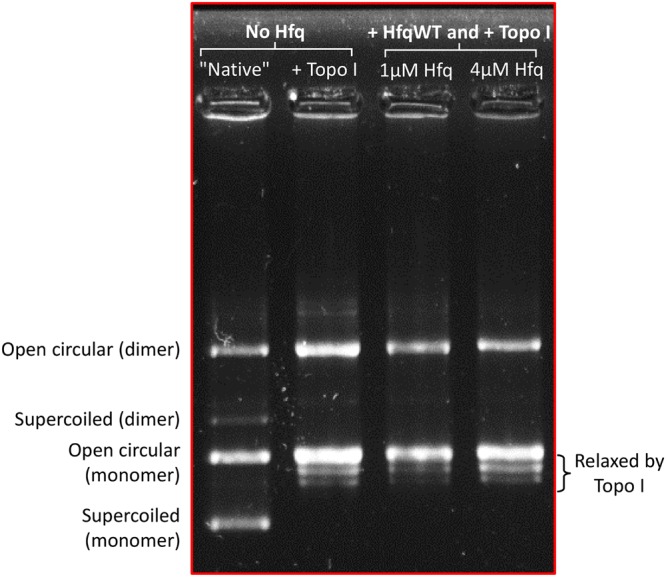
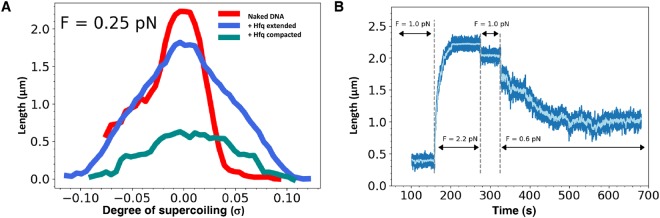
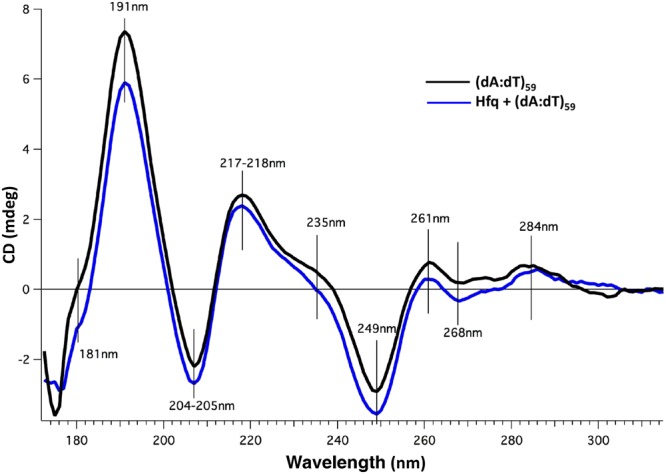
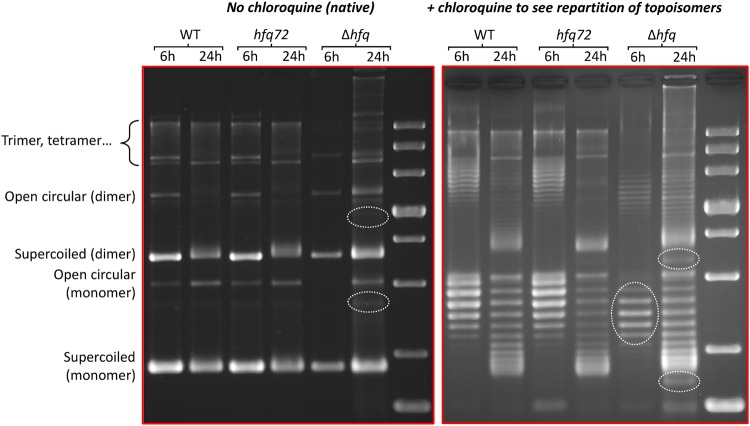
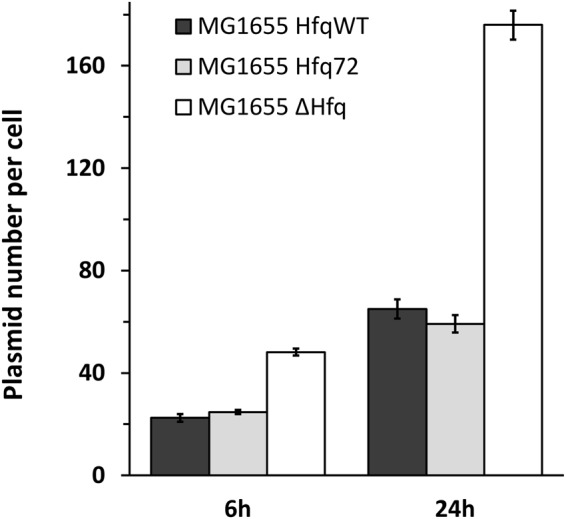
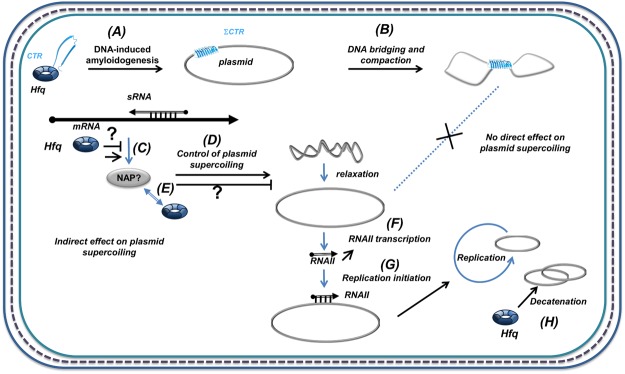
Hfq is a pleiotropic regulator that mediates several aspects of bacterial RNA metabolism. The protein notably regulates translation efficiency and RNA decay in Gram-negative bacteria, usually via its interaction with small regulatory RNA. Besides these RNA-related functions, Hfq has also been described as one of the nucleoid associated proteins shaping the bacterial chromosome. Therefore, Hfq appears as a versatile nucleic acid-binding protein, which functions are probably even more numerous than those initially suggested. For instance, E. coli Hfq, and more precisely its C-terminal region (CTR), has been shown to induce DNA compaction into a condensed form. In this paper, we establish that DNA induces Hfq-CTR amyloidogenesis, resulting in a change of DNA local conformation. Furthermore, we clarify the effect of Hfq on DNA topology. Our results evidence that, even if the protein has a strong propensity to compact DNA thanks to its amyloid region, it does not affect overall DNA topology. We confirm however that hfq gene disruption influences plasmid supercoiling in vivo, indicating that the effect on DNA topology in former reports was indirect. Most likely, this effect is related to small regulatory sRNA-Hfq-based regulation of another protein that influences DNA supercoiling, possibly a nucleoid associated protein such as H-NS or Dps. Finally, we hypothesise that this indirect effect on DNA topology explains, at least partially, the previously reported effect of Hfq on plasmid replication efficiency.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Franze de Fernandez MT, Hayward WS, August JT. Bacterial proteins required for replication of phage Qb ribonucleic acid. J. Biol. Chem. 1972;247:824–821. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
