

Single-cell reconstruction of the early maternal-fetal interface in humans
- PMID: 30429548
- PMCID: PMC7612850
- DOI: 10.1038/s41586-018-0698-6
Single-cell reconstruction of the early maternal-fetal interface in humans
Abstract
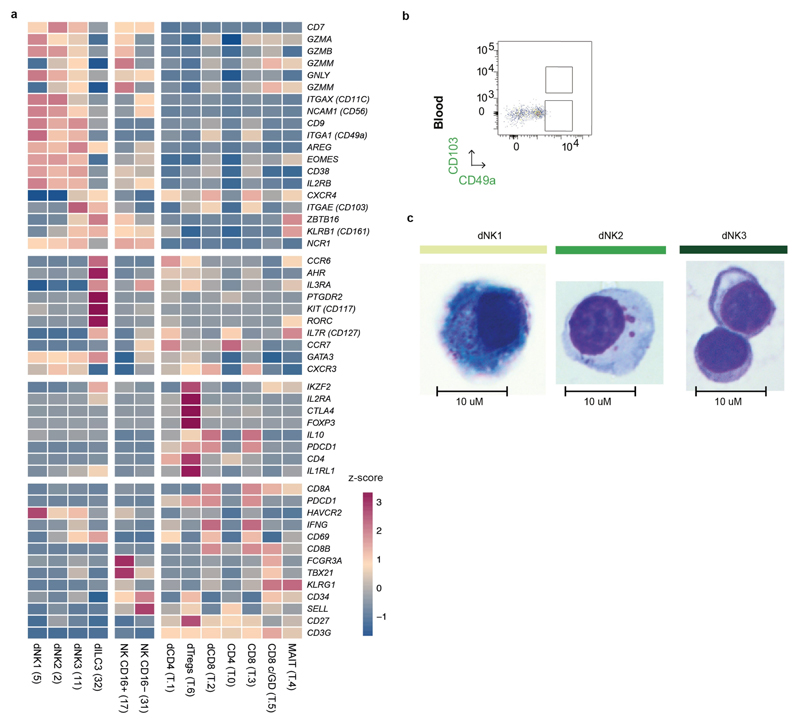
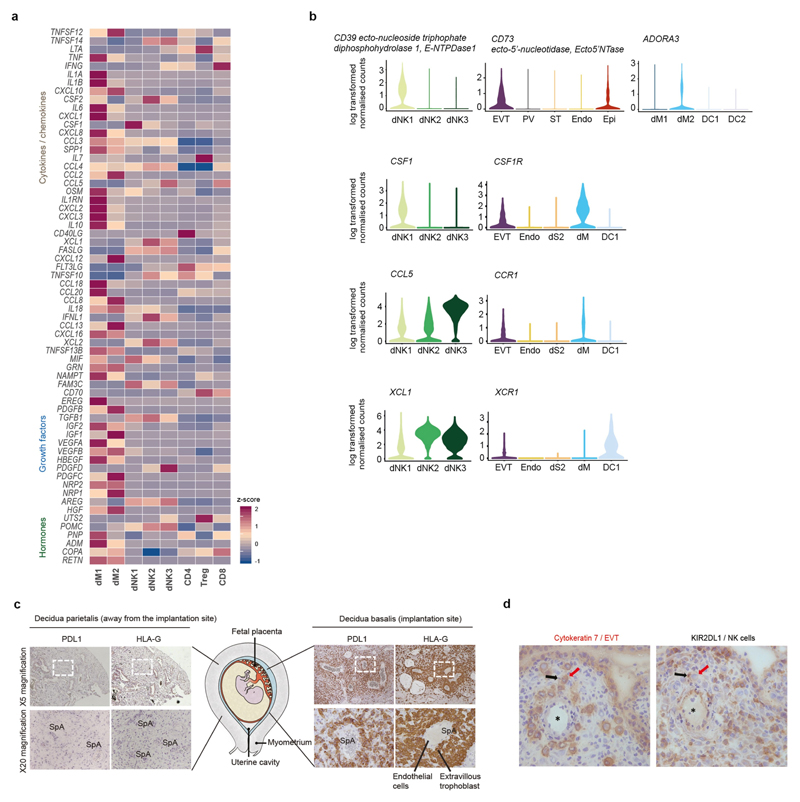
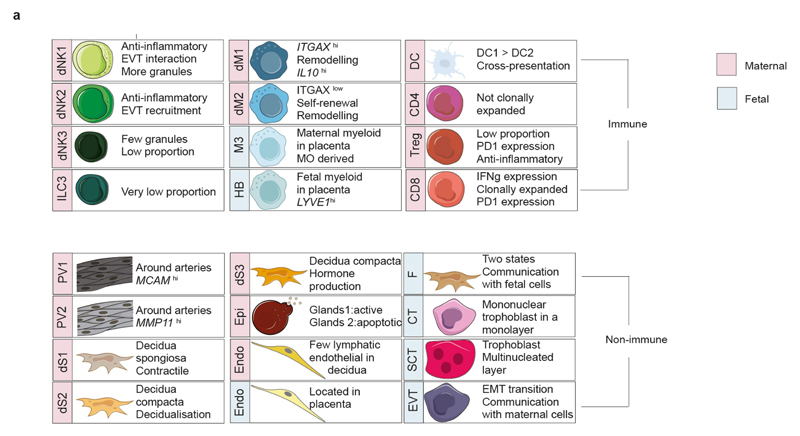
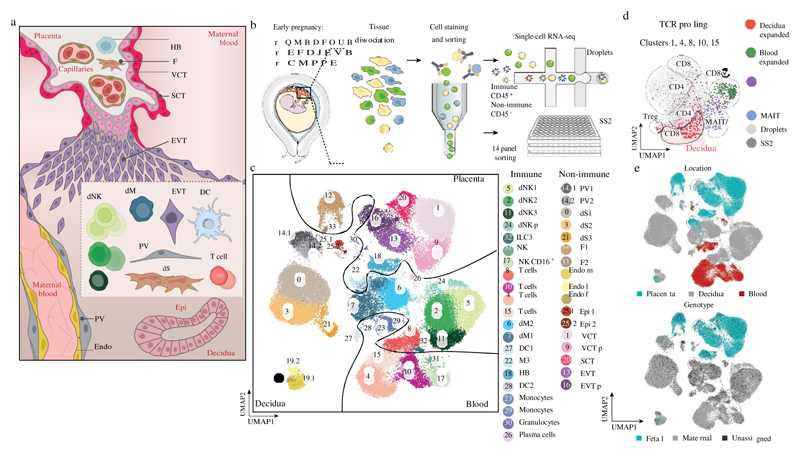
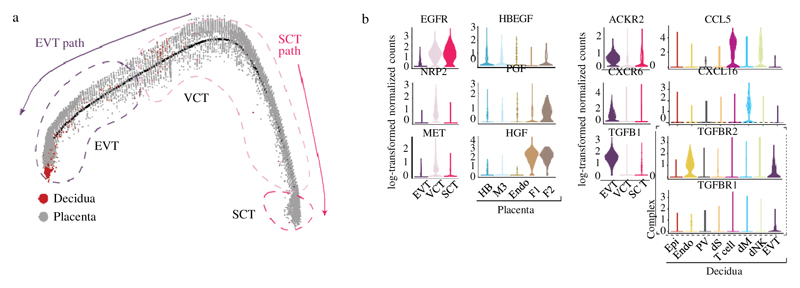
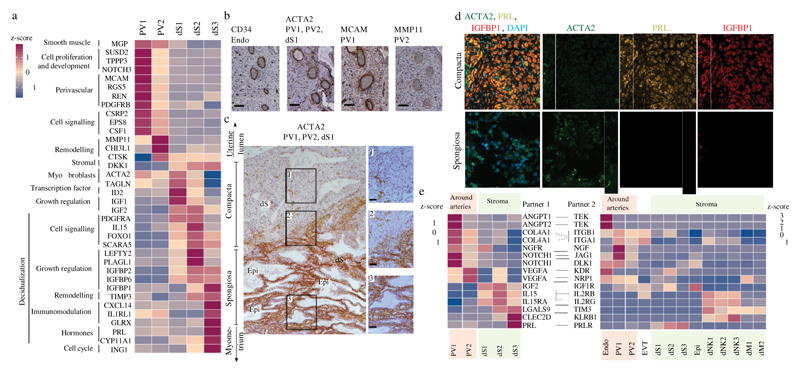
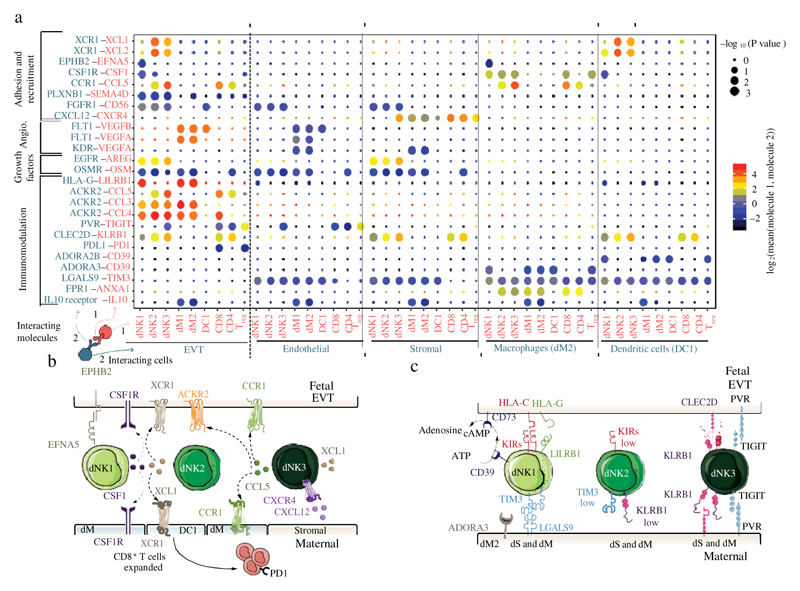
During early human pregnancy the uterine mucosa transforms into the decidua, into which the fetal placenta implants and where placental trophoblast cells intermingle and communicate with maternal cells. Trophoblast-decidual interactions underlie common diseases of pregnancy, including pre-eclampsia and stillbirth. Here we profile the transcriptomes of about 70,000 single cells from first-trimester placentas with matched maternal blood and decidual cells. The cellular composition of human decidua reveals subsets of perivascular and stromal cells that are located in distinct decidual layers. There are three major subsets of decidual natural killer cells that have distinctive immunomodulatory and chemokine profiles. We develop a repository of ligand-receptor complexes and a statistical tool to predict the cell-type specificity of cell-cell communication via these molecular interactions. Our data identify many regulatory interactions that prevent harmful innate or adaptive immune responses in this environment. Our single-cell atlas of the maternal-fetal interface reveals the cellular organization of the decidua and placenta, and the interactions that are critical for placentation and reproductive success.
Conflict of interest statement
Figures
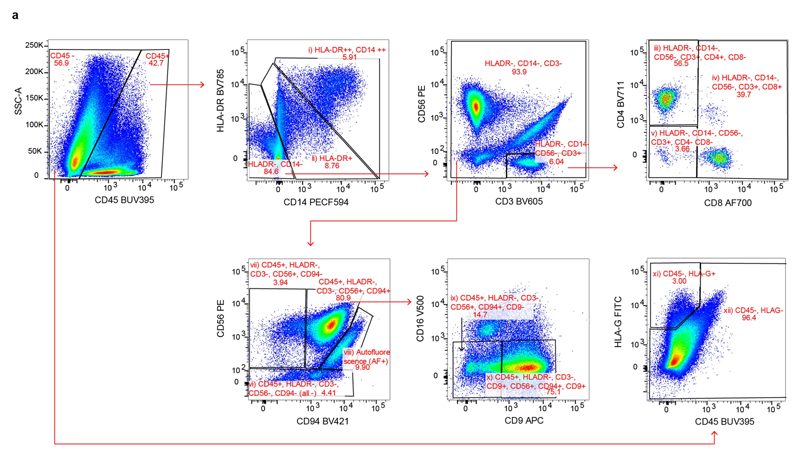

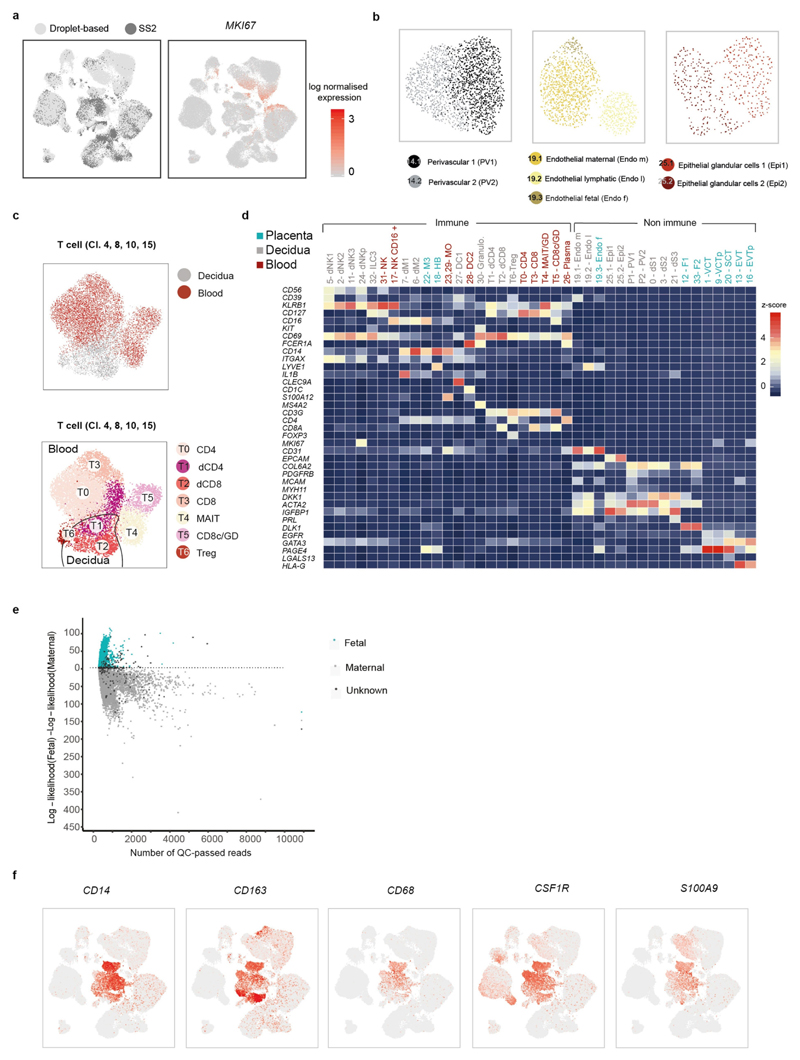
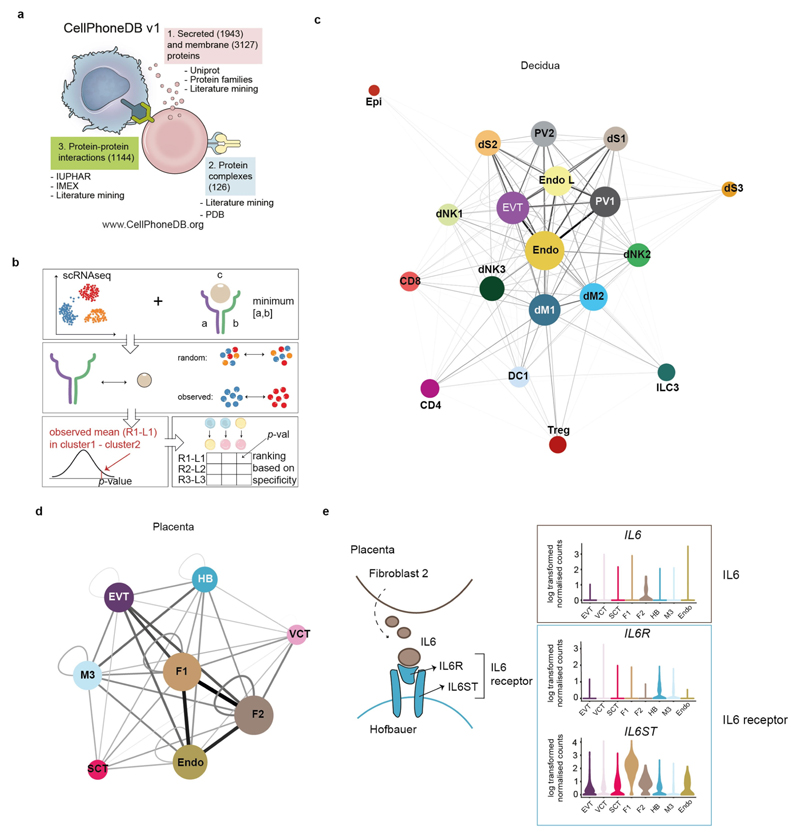
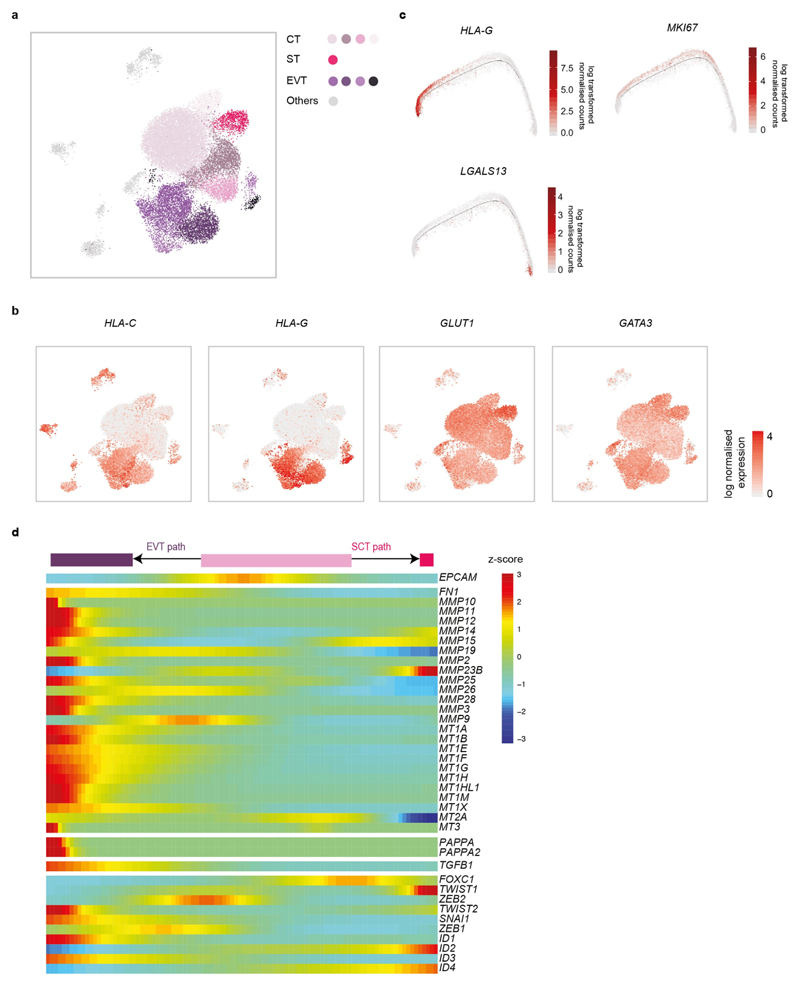



Comment in
-
Cell atlas reveals the landscape of early pregnancy.Nature. 2018 Nov;563(7731):337-338. doi: 10.1038/d41586-018-07317-w. Nature. 2018. PMID: 30429561 Free PMC article.
References
-
- Burton GJ, Watson AL, Hempstock J, Skepper JN, Jauniaux E. Uterine glands provide histiotrophic nutrition for the human fetus during the first trimester of pregnancy. J Clin Endocrinol Metab. 2002;87:2954–2959. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
