

Listeriolysin O-dependent host surfaceome remodeling modulates Listeria monocytogenes invasion
- PMID: 30445439
- PMCID: PMC6282100
- DOI: 10.1093/femspd/fty082
Listeriolysin O-dependent host surfaceome remodeling modulates Listeria monocytogenes invasion
Abstract
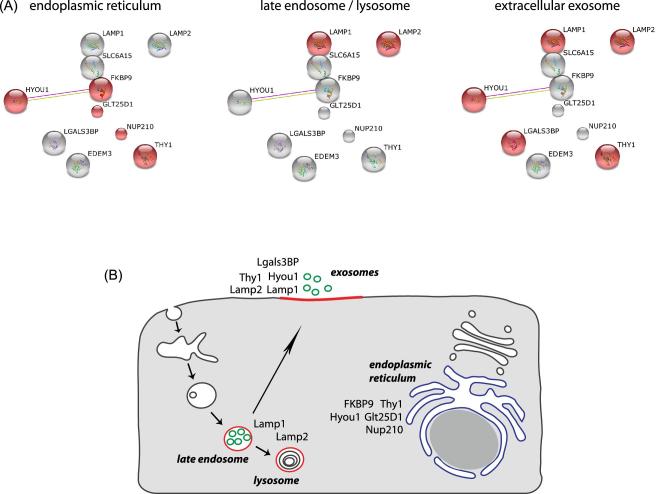
Listeria monocytogenes is a pathogenic bacterium that invades epithelial cells by activating host signaling cascades, which promote bacterial engulfment within a phagosome. The pore-forming toxin listeriolysin O (LLO), which is required for bacteria phagosomal escape, has also been associated with the activation of several signaling pathways when secreted by extracellular bacteria, including Ca2+ influx and promotion of L. monocytogenes entry. Quantitative host surfaceome analysis revealed significant quantitative remodeling of a defined set of cell surface glycoproteins upon LLO treatment, including a subset previously identified to play a role in the L. monocytogenes infection process. Our data further shows that the lysosomal-associated membrane proteins LAMP-1 and LAMP-2 are translocated to the cellular surface and those LLO-induced Ca2+ fluxes are required to trigger the surface relocalization of LAMP-1. Finally, we identify late endosomes/lysosomes as the major donor compartments of LAMP-1 upon LLO treatment and by perturbing their function, we suggest that these organelles participate in L. monocytogenes invasion.
Figures




Similar articles
-
The pore-forming toxin listeriolysin O mediates a novel entry pathway of L. monocytogenes into human hepatocytes.PLoS Pathog. 2011 Nov;7(11):e1002356. doi: 10.1371/journal.ppat.1002356. Epub 2011 Nov 3. PLoS Pathog. 2011. PMID: 22072970 Free PMC article.
-
Relative Roles of Listeriolysin O, InlA, and InlB in Listeria monocytogenes Uptake by Host Cells.Infect Immun. 2018 Sep 21;86(10):e00555-18. doi: 10.1128/IAI.00555-18. Print 2018 Oct. Infect Immun. 2018. PMID: 30061379 Free PMC article.
-
Listeriolysin O: a phagosome-specific lysin.Microbes Infect. 2007 Aug;9(10):1176-87. doi: 10.1016/j.micinf.2007.05.005. Epub 2007 May 7. Microbes Infect. 2007. PMID: 17720603 Review.
-
Fluxes of Ca2+ and K+ are required for the listeriolysin O-dependent internalization pathway of Listeria monocytogenes.Infect Immun. 2014 Mar;82(3):1084-91. doi: 10.1128/IAI.01067-13. Epub 2013 Dec 23. Infect Immun. 2014. PMID: 24366251 Free PMC article.
-
Listeriolysin O: A phagosome-specific cytolysin revisited.Cell Microbiol. 2019 Mar;21(3):e12988. doi: 10.1111/cmi.12988. Epub 2019 Jan 15. Cell Microbiol. 2019. PMID: 30511471 Free PMC article. Review.
Cited by
-
Eeukaryotic-like Sppsk1 from Vibrio splendidus AJ01 mediates phagosome escape via inhibiting phagosome acidification and maturation.Cell Mol Life Sci. 2025 Feb 22;82(1):88. doi: 10.1007/s00018-025-05610-2. Cell Mol Life Sci. 2025. PMID: 39985586 Free PMC article.
-
Toxins, Pathogenicity, Anti-Toxins, a Bicentennial Contribution.Toxins (Basel). 2024 Feb 10;16(2):97. doi: 10.3390/toxins16020097. Toxins (Basel). 2024. PMID: 38393174 Free PMC article.
-
Infectious Threats, the Intestinal Barrier, and Its Trojan Horse: Dysbiosis.Front Microbiol. 2019 Aug 7;10:1676. doi: 10.3389/fmicb.2019.01676. eCollection 2019. Front Microbiol. 2019. PMID: 31447793 Free PMC article. Review.
-
Comparison of genetic variations between high- and low-risk Listeria monocytogenes isolates using whole-genome de novo sequencing.Front Microbiol. 2023 Jul 18;14:1163841. doi: 10.3389/fmicb.2023.1163841. eCollection 2023. Front Microbiol. 2023. PMID: 37533826 Free PMC article.
-
Cronobacter sakazakii ATCC 29544 Translocated Human Brain Microvascular Endothelial Cells via Endocytosis, Apoptosis Induction, and Disruption of Tight Junction.Front Microbiol. 2021 Jun 7;12:675020. doi: 10.3389/fmicb.2021.675020. eCollection 2021. Front Microbiol. 2021. PMID: 34163451 Free PMC article.
References
-
- Bierne H, Miki H, Innocenti M et al. . WASP-related proteins, Abi1 and Ena/VASP are required for Listeria invasion induced by the Met receptor. J Cell Sci 2005;118:1537–47. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
