

Aryl Hydrocarbon Receptor Signaling Cell Intrinsically Inhibits Intestinal Group 2 Innate Lymphoid Cell Function
- PMID: 30446384
- PMCID: PMC6249058
- DOI: 10.1016/j.immuni.2018.09.015
Aryl Hydrocarbon Receptor Signaling Cell Intrinsically Inhibits Intestinal Group 2 Innate Lymphoid Cell Function
Abstract
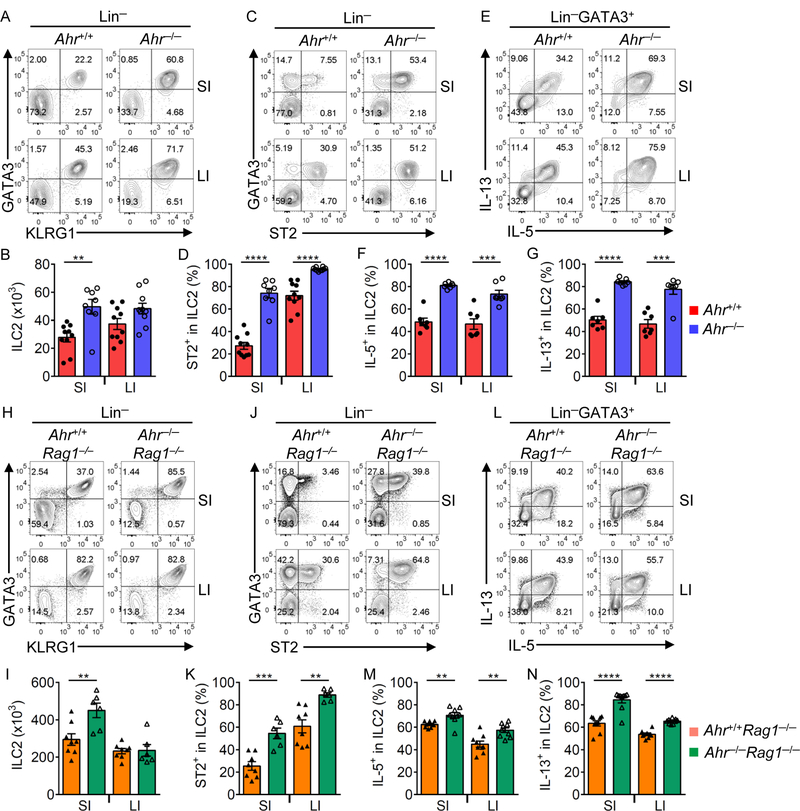
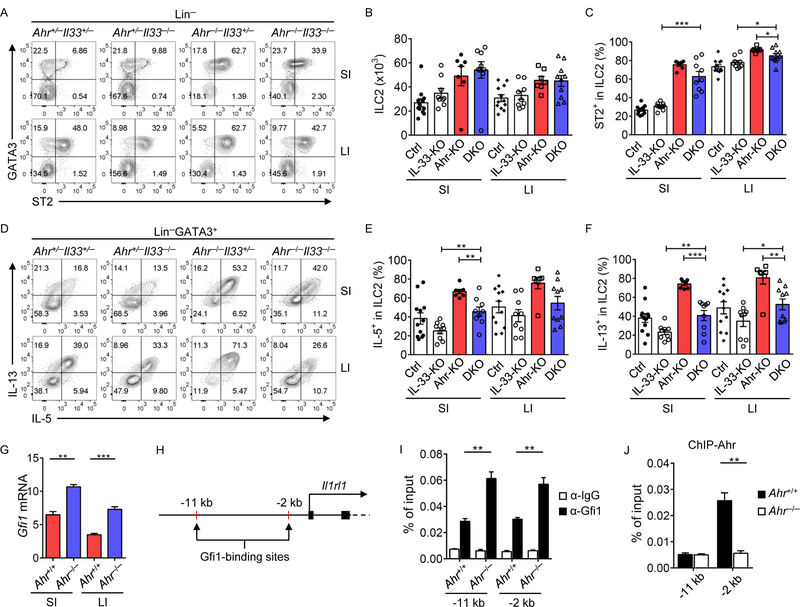
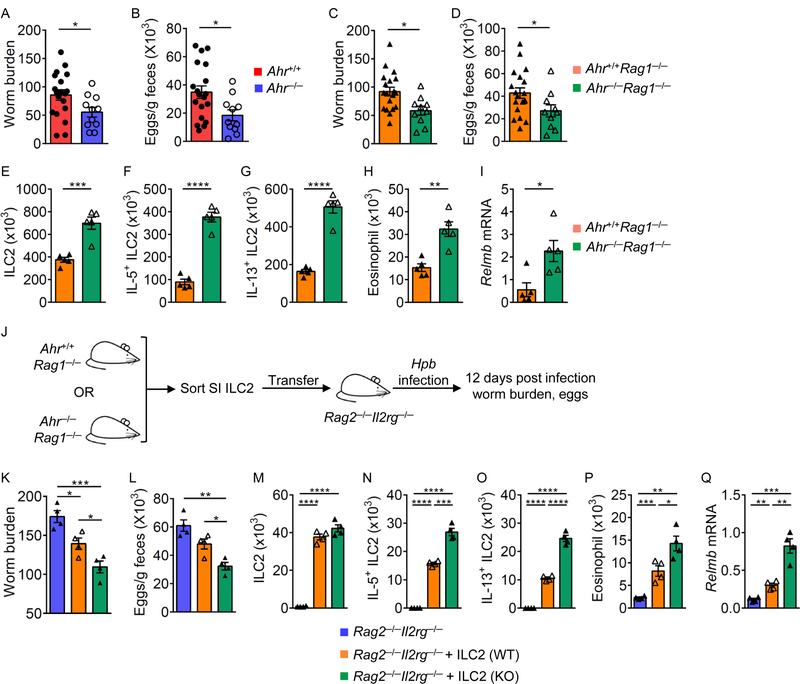
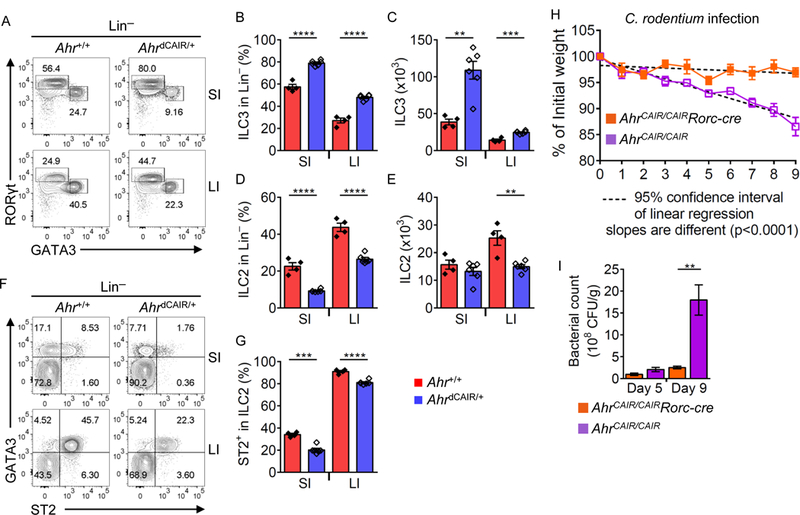
Innate lymphoid cells (ILCs) are important for mucosal immunity. The intestine harbors all ILC subsets, but how these cells are balanced to achieve immune homeostasis and mount appropriate responses during infection remains elusive. Here, we show that aryl hydrocarbon receptor (Ahr) expression in the gut regulates ILC balance. Among ILCs, Ahr is most highly expressed by gut ILC2s and controls chromatin accessibility at the Ahr locus via positive feedback. Ahr signaling suppresses Gfi1 transcription-factor-mediated expression of the interleukin-33 (IL-33) receptor ST2 in ILC2s and expression of ILC2 effector molecules IL-5, IL-13, and amphiregulin in a cell-intrinsic manner. Ablation of Ahr enhances anti-helminth immunity in the gut, whereas genetic or pharmacological activation of Ahr suppresses ILC2 function but enhances ILC3 maintenance to protect the host from Citrobacter rodentium infection. Thus, the host regulates the gut ILC2-ILC3 balance by engaging the Ahr pathway to mount appropriate immunity against various pathogens.
Keywords: Ahr; IL-33; ILC; ST2; chromatin; gene expression; gut immunity.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests
The authors declare no competing interests.
Figures



References
-
- Artis D, and Spits H (2015). The biology of innate lymphoid cells. Nature 517, 293–301. - PubMed
-
- Artis D, Wang ML, Keilbaugh SA, He W, Brenes M, Swain GP, Knight PA, Donaldson DD, Lazar MA, Miller HR, et al. (2004). RELMbeta/FIZZ2 is a goblet cell-specific immune-effector molecule in the gastrointestinal tract. Proceedings of the National Academy of Sciences of the United States of America 101, 13596–13600. - PMC - PubMed
-
- Bjorklund AK, Forkel M, Picelli S, Konya V, Theorell J, Friberg D, Sandberg R, and Mjosberg J (2016). The heterogeneity of human CD127(+) innate lymphoid cells revealed by single-cell RNA sequencing. Nat Immunol 17, 451–460. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
