

Spatiotemporal control of axillary meristem formation by interacting transcriptional regulators
- PMID: 30446629
- PMCID: PMC6307885
- DOI: 10.1242/dev.158352
Spatiotemporal control of axillary meristem formation by interacting transcriptional regulators
Abstract
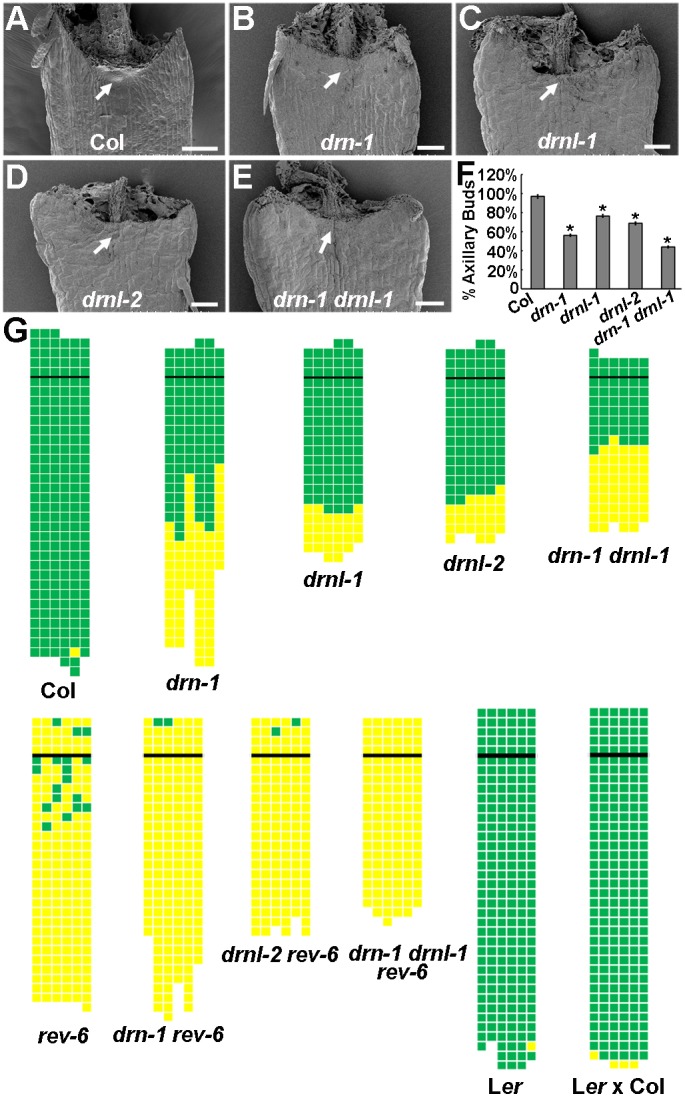
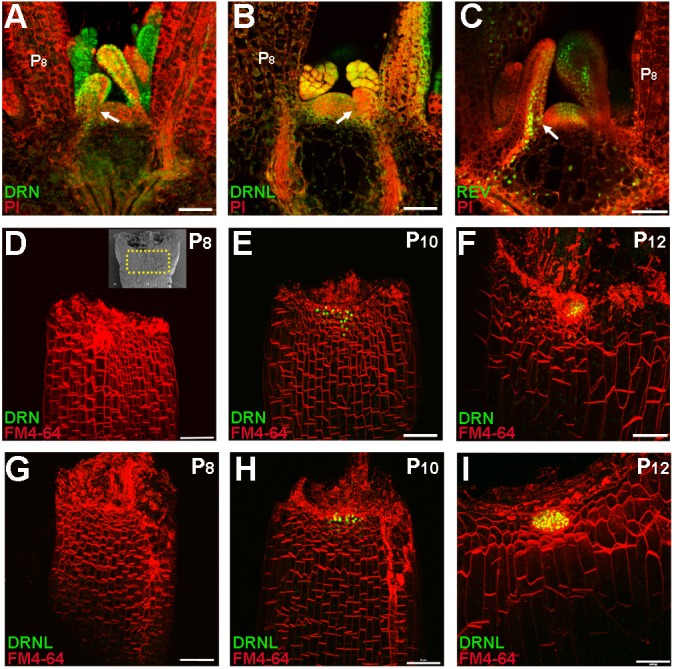
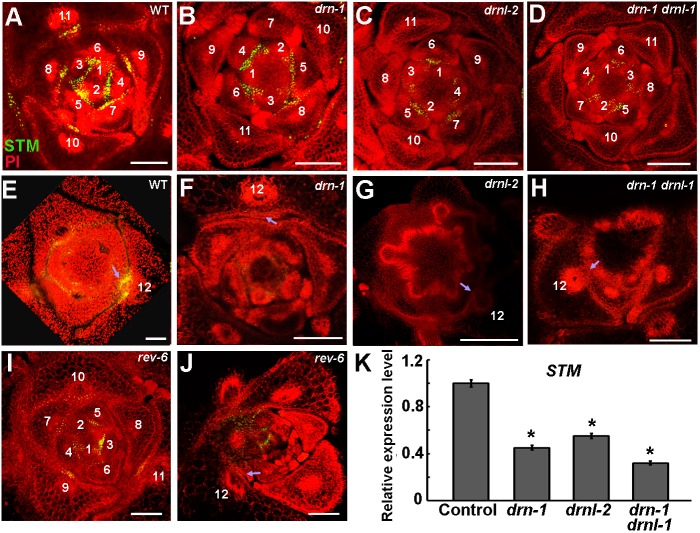
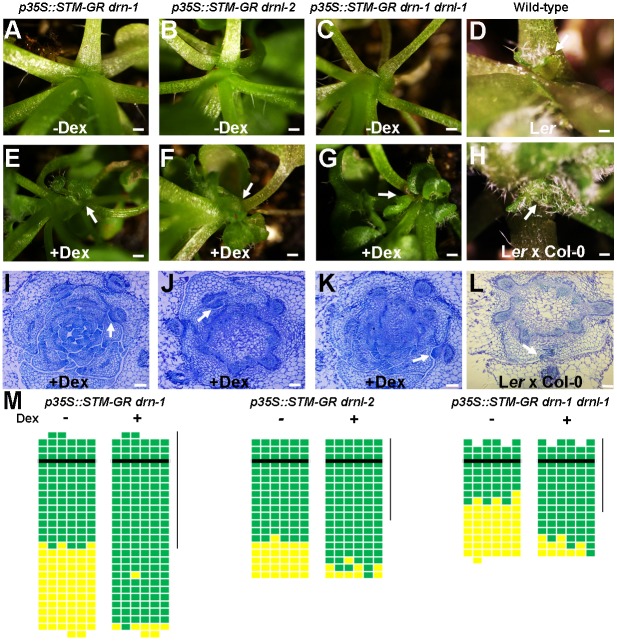
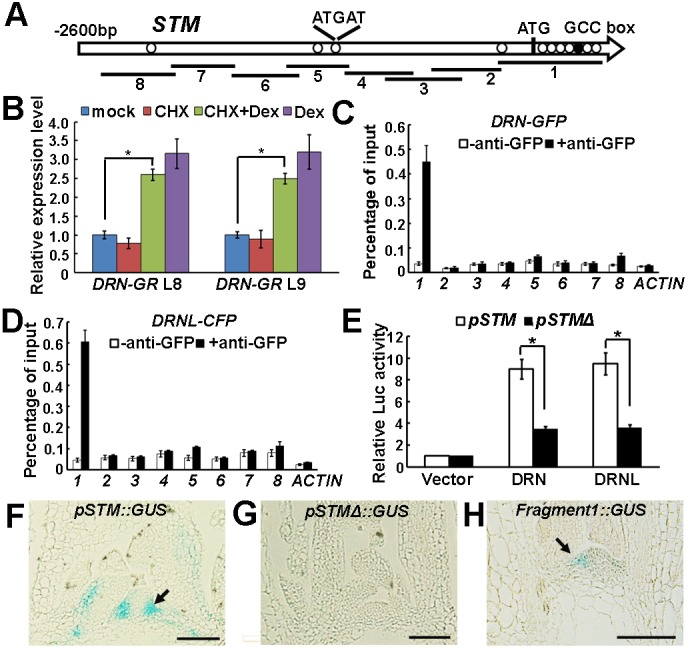
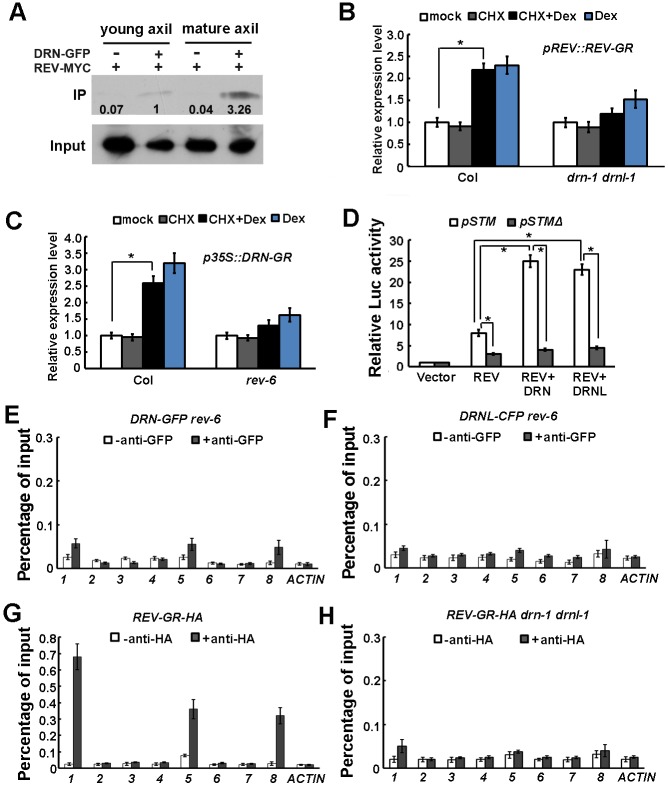
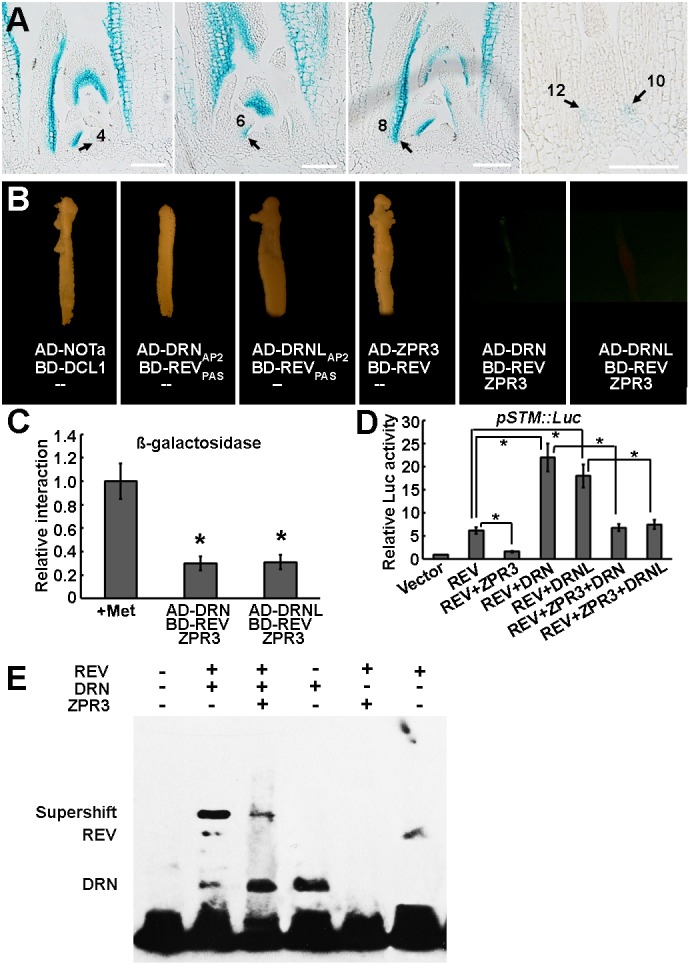
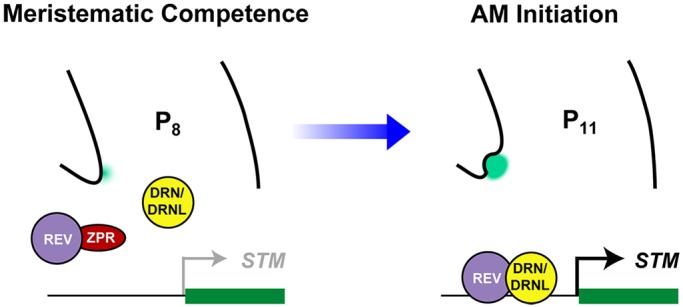
Branching is a common feature of plant development. In seed plants, axillary meristems (AMs) initiate in leaf axils to enable lateral shoot branching. AM initiation requires a high level of expression of the meristem marker SHOOT MERISTEMLESS (STM) in the leaf axil. Here, we show that modules of interacting transcriptional regulators control STM expression and AM initiation. Two redundant AP2-type transcription factors, DORNRÖSCHEN (DRN) and DORNRÖSCHEN-LIKE (DRNL), control AM initiation by regulating STM expression. DRN and DRNL directly upregulate STM expression in leaf axil meristematic cells, as does another transcription factor, REVOLUTA (REV). The activation of STM expression by DRN/DRNL depends on REV, and vice versa. DRN/DRNL and REV have overlapping expression patterns and protein interactions in the leaf axil, which are required for the upregulation of STM expression. Furthermore, LITTLE ZIPPER3, another REV-interacting protein, is expressed in the leaf axil and interferes with the DRN/DRNL-REV interaction to negatively modulate STM expression. Our results support a model in which interacting transcriptional regulators fine-tune the expression of STM to precisely regulate AM initiation. Thus, shoot branching recruits the same conserved protein complexes used in embryogenesis and leaf polarity patterning.
Keywords: Arabidopsis; Axillary meristem; Branching; Stem cell; Transcription.
© 2018. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
References
-
- Aguilar-Martínez J. A., Uchida N., Townsley B., West D. A., Yanez A., Lynn N., Kimura S. and Sinha N. (2015). Transcriptional, posttranscriptional, and posttranslational regulation of SHOOT MERISTEMLESS gene expression in Arabidopsis determines gene function in the shoot apex. Plant Physiol. 167, 424-442. 10.1104/pp.114.248625 - DOI - PMC - PubMed
-
- Banno H., Mase H. and Maekawa K. (2006). Analysis of functional domains and binding sequences of Arabidopsis transcription factor ESR1. Plant Biotechnol. 23, 303-308. 10.5511/plantbiotechnology.23.303 - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
