

Dna2 processes behind the fork long ssDNA flaps generated by Pif1 and replication-dependent strand displacement
- PMID: 30446656
- PMCID: PMC6240037
- DOI: 10.1038/s41467-018-07378-5
Dna2 processes behind the fork long ssDNA flaps generated by Pif1 and replication-dependent strand displacement
Abstract
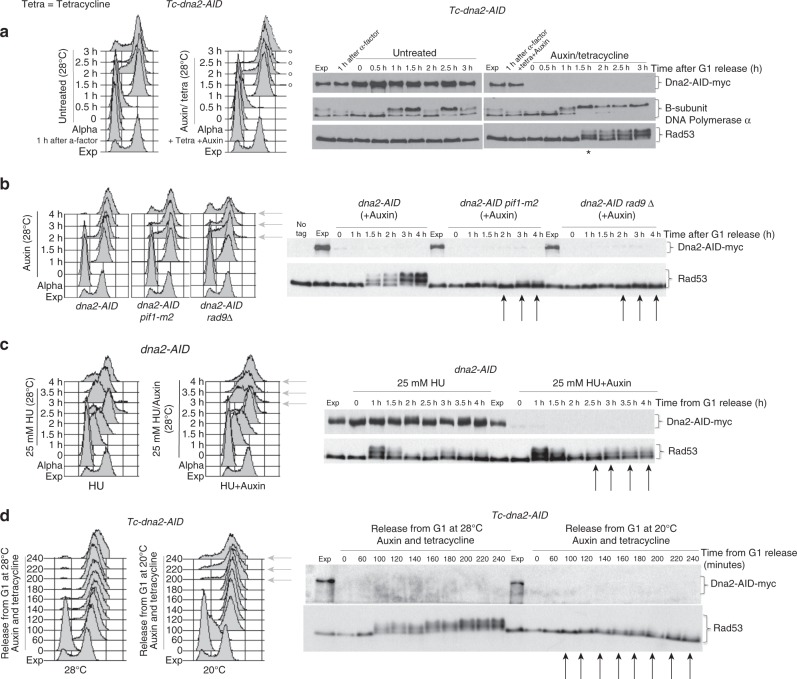
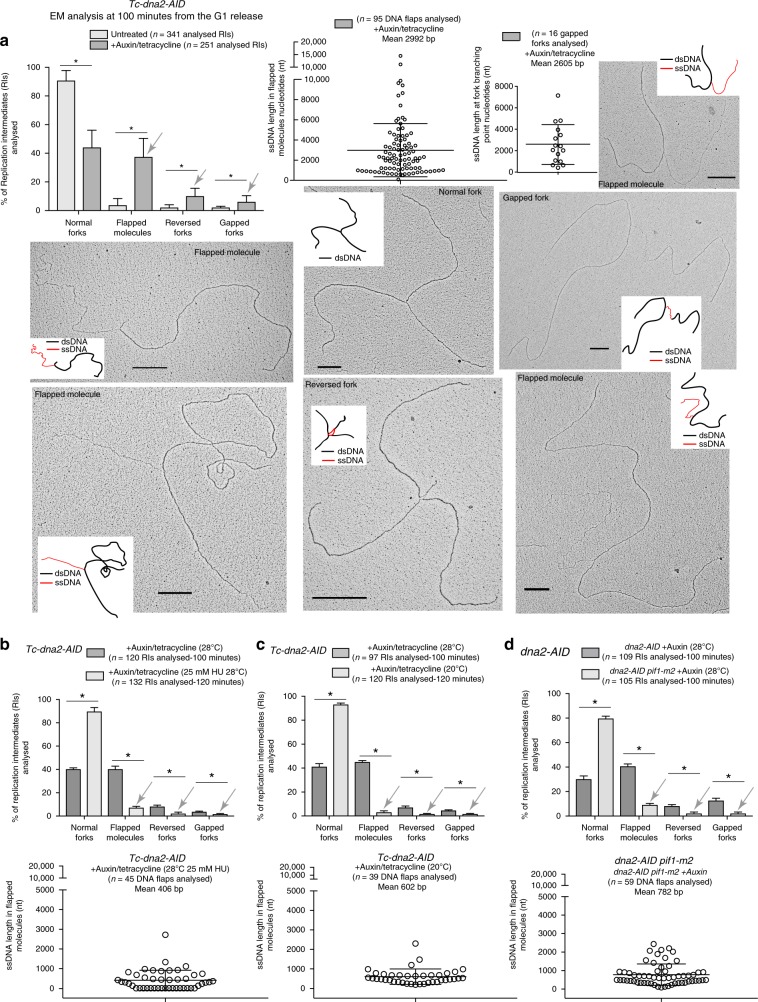
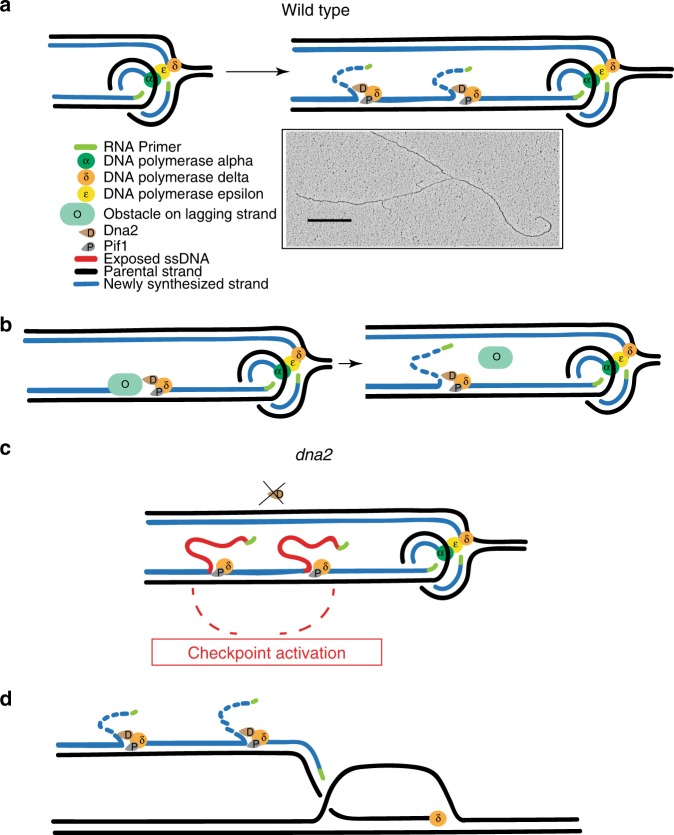
Dna2 is a DNA helicase-endonuclease mediating DSB resection and Okazaki fragment processing. Dna2 ablation is lethal and rescued by inactivation of Pif1, a helicase assisting Okazaki fragment maturation, Pol32, a DNA polymerase δ subunit, and Rad9, a DNA damage response (DDR) factor. Dna2 counteracts fork reversal and promotes fork restart. Here we show that Dna2 depletion generates lethal DNA structures activating the DDR. While PIF1 deletion rescues the lethality of Dna2 depletion, RAD9 ablation relieves the first cell cycle arrest causing genotoxicity after few cell divisions. Slow fork speed attenuates DDR in Dna2 deprived cells. Electron microscopy shows that Dna2-ablated cells accumulate long ssDNA flaps behind the forks through Pif1 and fork speed. We suggest that Dna2 offsets the strand displacement activity mediated by the lagging strand polymerase and Pif1, processing long ssDNA flaps to prevent DDR activation. We propose that this Dna2 function has been hijacked by Break Induced Replication in DSB processing.
Conflict of interest statement
The authors declare no competing interests.
Figures


Similar articles
-
Disease-associated DNA2 nuclease-helicase protects cells from lethal chromosome under-replication.Nucleic Acids Res. 2020 Jul 27;48(13):7265-7278. doi: 10.1093/nar/gkaa524. Nucleic Acids Res. 2020. PMID: 32544229 Free PMC article.
-
Evidence suggesting that Pif1 helicase functions in DNA replication with the Dna2 helicase/nuclease and DNA polymerase delta.Mol Cell Biol. 2006 Apr;26(7):2490-500. doi: 10.1128/MCB.26.7.2490-2500.2006. Mol Cell Biol. 2006. PMID: 16537895 Free PMC article.
-
An alternative pathway for Okazaki fragment processing: resolution of fold-back flaps by Pif1 helicase.J Biol Chem. 2010 Dec 31;285(53):41712-23. doi: 10.1074/jbc.M110.146894. Epub 2010 Oct 19. J Biol Chem. 2010. PMID: 20959454 Free PMC article.
-
Yet another job for Dna2: Checkpoint activation.DNA Repair (Amst). 2015 Aug;32:17-23. doi: 10.1016/j.dnarep.2015.04.009. Epub 2015 May 1. DNA Repair (Amst). 2015. PMID: 25956863 Free PMC article. Review.
-
DNA2-An Important Player in DNA Damage Response or Just Another DNA Maintenance Protein?Int J Mol Sci. 2017 Jul 18;18(7):1562. doi: 10.3390/ijms18071562. Int J Mol Sci. 2017. PMID: 28718810 Free PMC article. Review.
Cited by
-
Control of Replication Stress Response by Cytosolic Fe-S Cluster Assembly (CIA) Machinery.Cells. 2025 Mar 16;14(6):442. doi: 10.3390/cells14060442. Cells. 2025. PMID: 40136691 Free PMC article. Review.
-
RPA and Pif1 cooperate to remove G-rich structures at both leading and lagging strand.Cell Stress. 2020 Jan 17;4(3):48-63. doi: 10.15698/cst2020.03.214. Cell Stress. 2020. PMID: 32190820 Free PMC article.
-
Limiting homologous recombination at stalled replication forks is essential for cell viability: DNA2 to the rescue.Curr Genet. 2020 Dec;66(6):1085-1092. doi: 10.1007/s00294-020-01106-7. Epub 2020 Sep 9. Curr Genet. 2020. PMID: 32909097 Free PMC article. Review.
-
Pif1, RPA, and FEN1 modulate the ability of DNA polymerase δ to overcome protein barriers during DNA synthesis.J Biol Chem. 2020 Nov 20;295(47):15883-15891. doi: 10.1074/jbc.RA120.015699. Epub 2020 Sep 10. J Biol Chem. 2020. PMID: 32913126 Free PMC article.
-
Post-replicative nick translation occurs on the lagging strand during prolonged depletion of DNA ligase I in Saccharomyces cerevisiae.G3 (Bethesda). 2021 Aug 7;11(8):jkab205. doi: 10.1093/g3journal/jkab205. G3 (Bethesda). 2021. PMID: 34849819 Free PMC article.
References
-
- Maga G, et al. Okazaki fragment processing: modulation of the strand displacement activity of DNA polymerase delta by the concerted action of replication protein A, proliferating cell nuclear antigen, and flap endonuclease-1. Proc. Natl. Acad. Sci. USA. 2001;98:14298–14303. doi: 10.1073/pnas.251193198. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
