

Microtubule Feedback and LET-99-Dependent Control of Pulling Forces Ensure Robust Spindle Position
- PMID: 30447992
- PMCID: PMC6289040
- DOI: 10.1016/j.bpj.2018.10.010
Microtubule Feedback and LET-99-Dependent Control of Pulling Forces Ensure Robust Spindle Position
Abstract
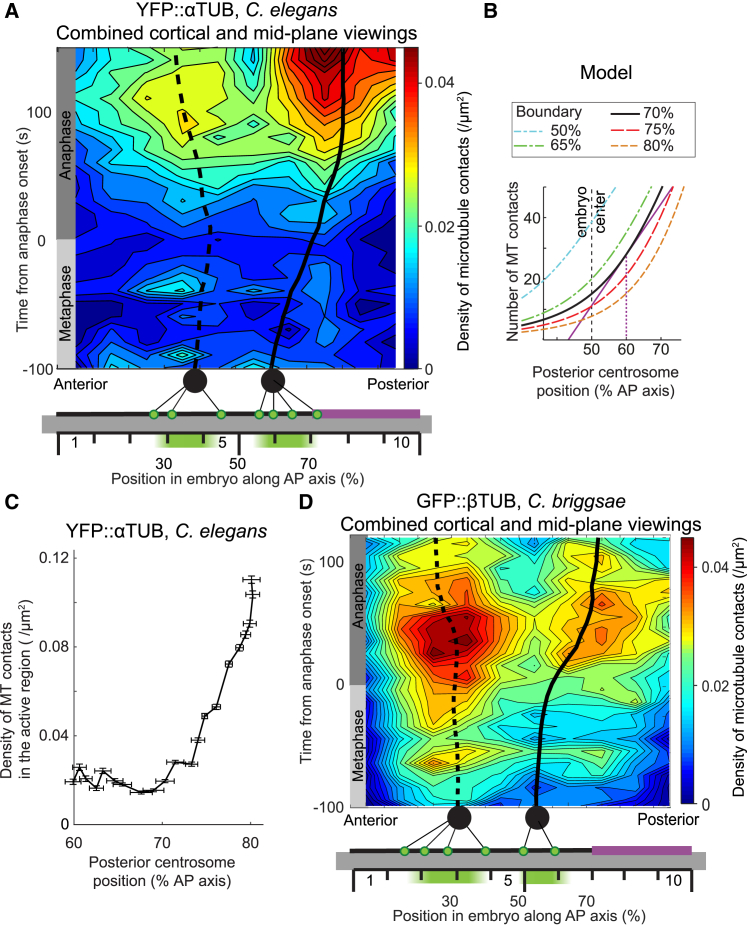
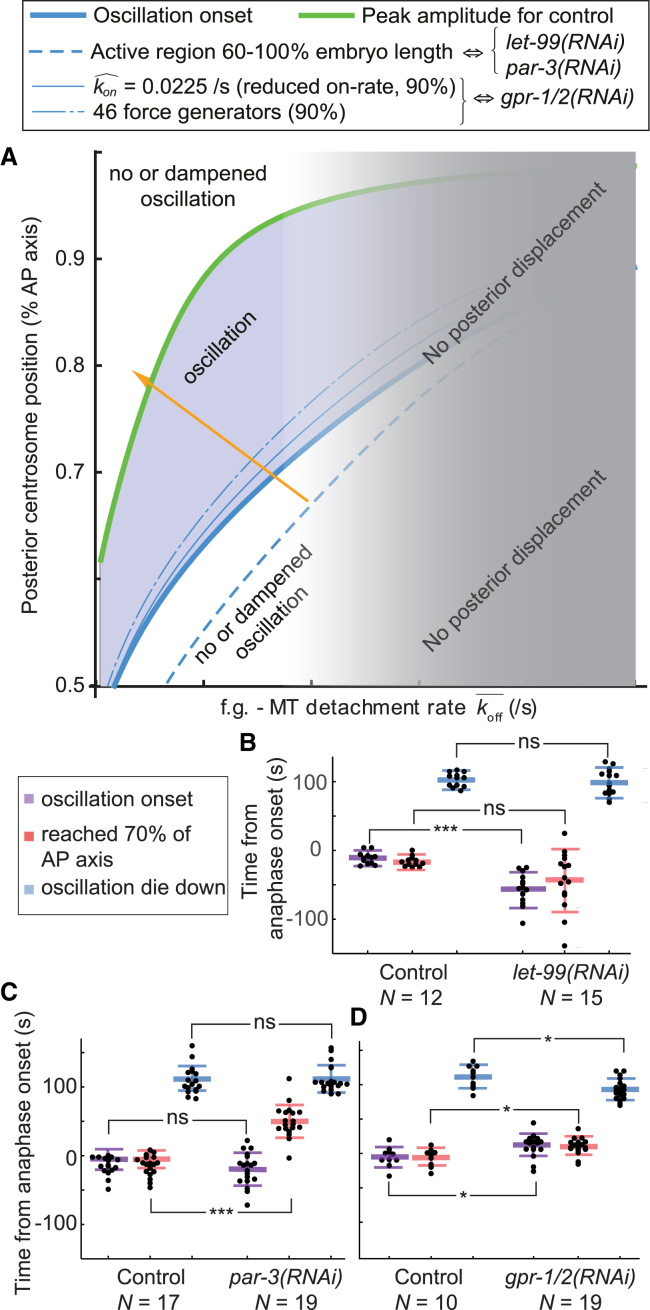
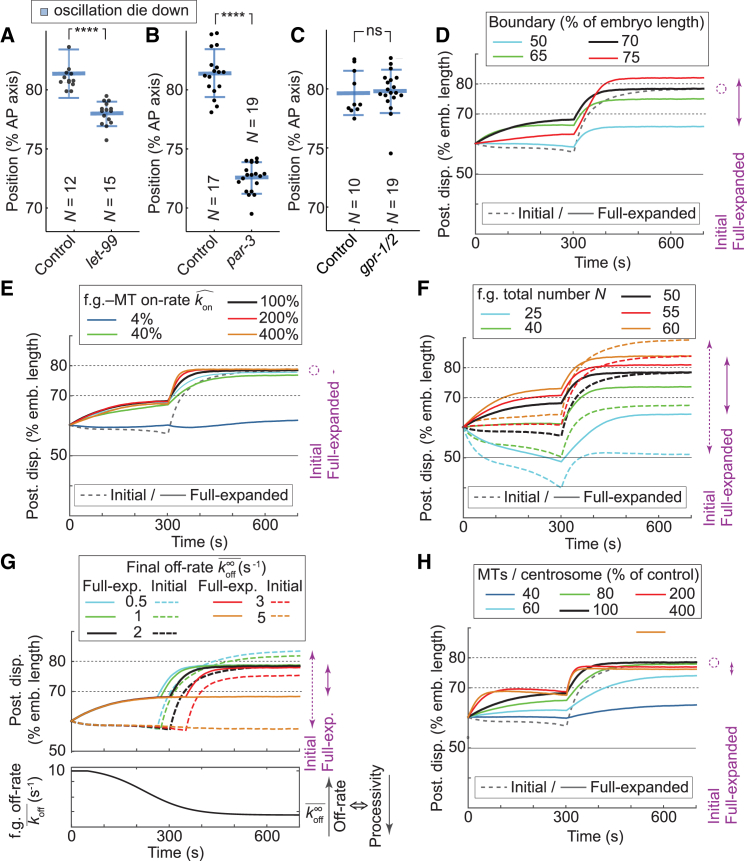
During asymmetric division of the Caenorhabditis elegans zygote, to properly distribute cell fate determinants, the mitotic spindle is asymmetrically localized by a combination of centering and cortical-pulling microtubule-mediated forces, the dynamics of the latter being regulated by mitotic progression. Here, we show a, to our knowledge, novel and additional regulation of these forces by spindle position itself. For that, we observed the onset of transverse spindle oscillations, which reflects the burst of anaphase pulling forces. After delaying anaphase onset, we found that the position at which the spindle starts to oscillate was unchanged compared to control embryos and uncorrelated to anaphase onset. In mapping the cortical microtubule dynamics, we measured a steep increase in microtubule contact density after the posterior centrosome reached the critical position of 70% of embryo length, strongly suggesting the presence of a positional switch for spindle oscillations. Expanding a previous model based on a force-generator temporal control, we implemented this positional switch and observed that the large increase in microtubule density accounted for the pulling force burst. Thus, we propose that the spindle position influences the cortical availability of microtubules on which the active force generators, controlled by cell cycle progression, can pull. Importantly, we found that this positional control relies on the polarity-dependent LET-99 cortical band, the boundary of which could be probed by microtubules. This dual positional and temporal control well accounted for our observation that the oscillation onset position resists changes in cellular geometry and moderate variations in the active force generator number. Finally, our model suggests that spindle position at mitosis end is more sensitive to the polarity factor LET-99, which restricts the region of active force generators to a posterior-most region, than to microtubule number or force generator number/activity. Overall, we show that robustness in spindle positioning originates in cell mechanics rather than biochemical networks.
Copyright © 2018 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures



References
-
- Betschinger J., Knoblich J.A. Dare to be different: asymmetric cell division in Drosophila, C. elegans and vertebrates. Curr. Biol. 2004;14:R674–R685. - PubMed
-
- Gillies T.E., Cabernard C. Cell division orientation in animals. Curr. Biol. 2011;21:R599–R609. - PubMed
-
- Gönczy P. Mechanisms of asymmetric cell division: flies and worms pave the way. Nat. Rev. Mol. Cell Biol. 2008;9:355–366. - PubMed
-
- Morin X., Bellaïche Y. Mitotic spindle orientation in asymmetric and symmetric cell divisions during animal development. Dev. Cell. 2011;21:102–119. - PubMed
