

RETRACTED: Peptide-induced formation of protein aggregates and amyloid fibrils in human and guinea pig αA-crystallins under physiological conditions of temperature and pH
- PMID: 30448341
- PMCID: PMC6380173
- DOI: 10.1016/j.exer.2018.11.016
RETRACTED: Peptide-induced formation of protein aggregates and amyloid fibrils in human and guinea pig αA-crystallins under physiological conditions of temperature and pH
Retraction in
-
Retraction notice to "Peptide-induced formation of protein aggregates and amyloid fibrils in human and Guinea pig αA-crystallins under physiological conditions of temperature and pH" [Exp. Eye Res. 179 (2019) 193-205].Exp Eye Res. 2020 May;194:107990. doi: 10.1016/j.exer.2020.107990. Epub 2020 Apr 16. Exp Eye Res. 2020. PMID: 32307097 Free PMC article.
Abstract
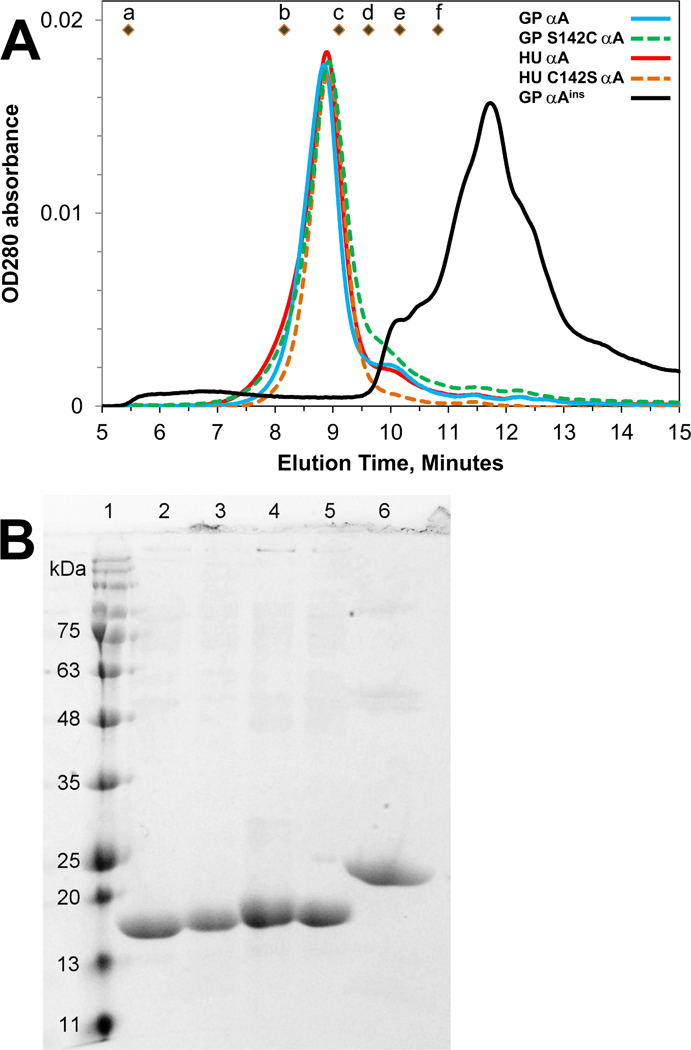
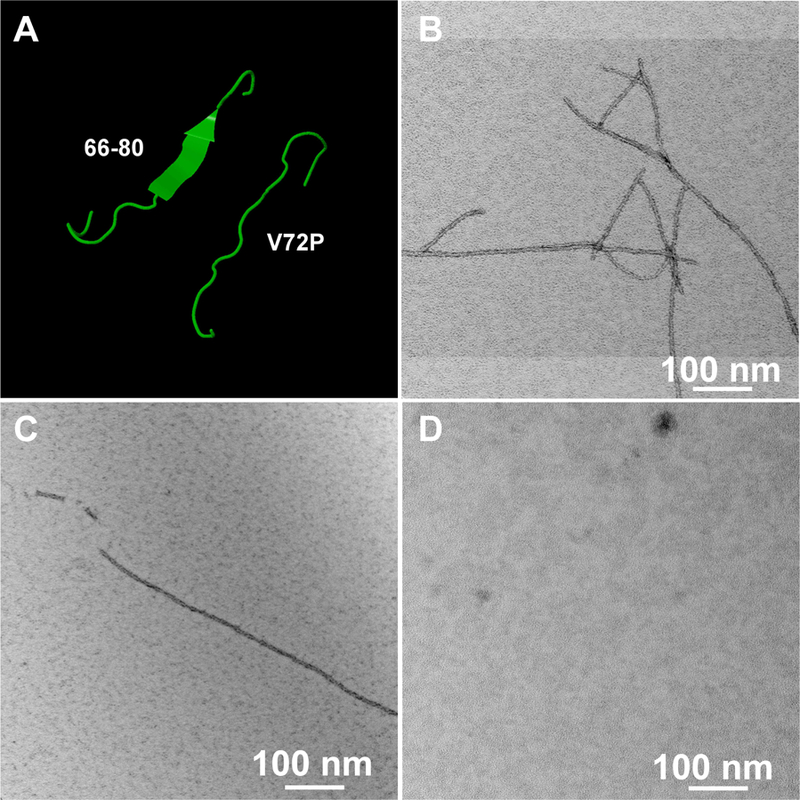
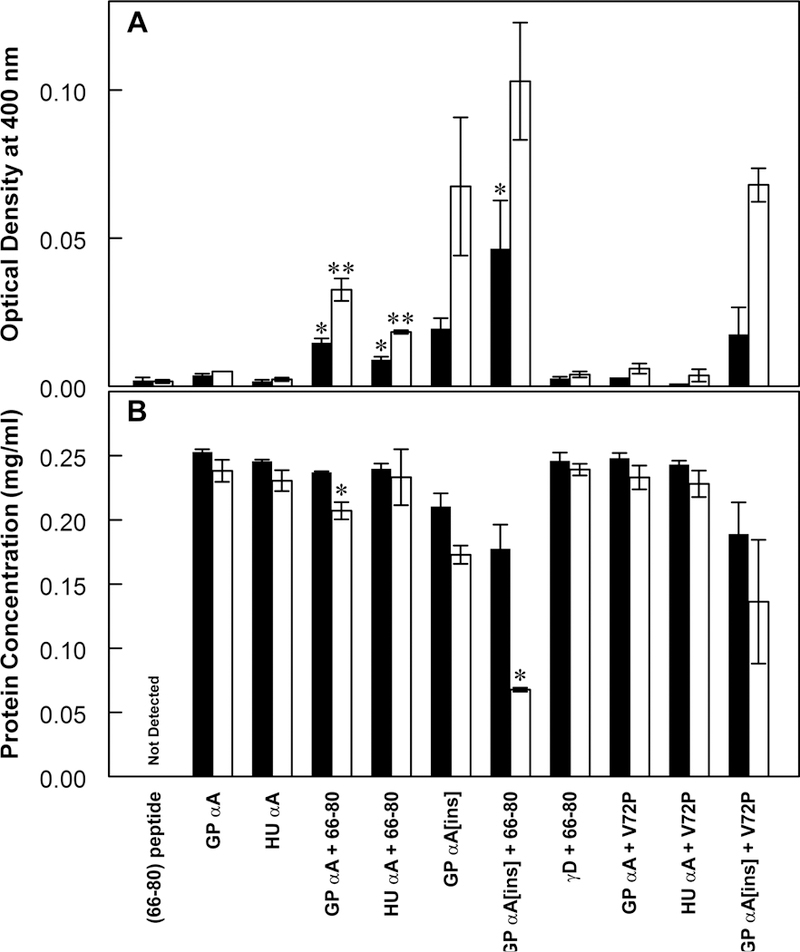
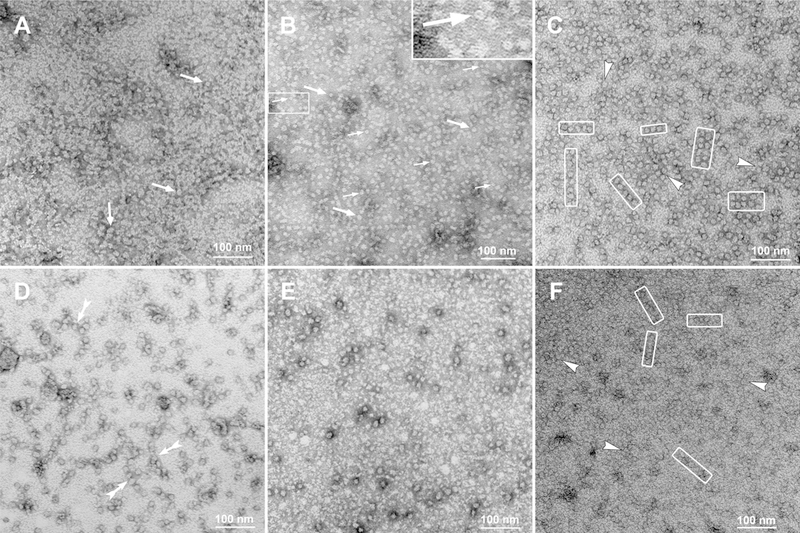
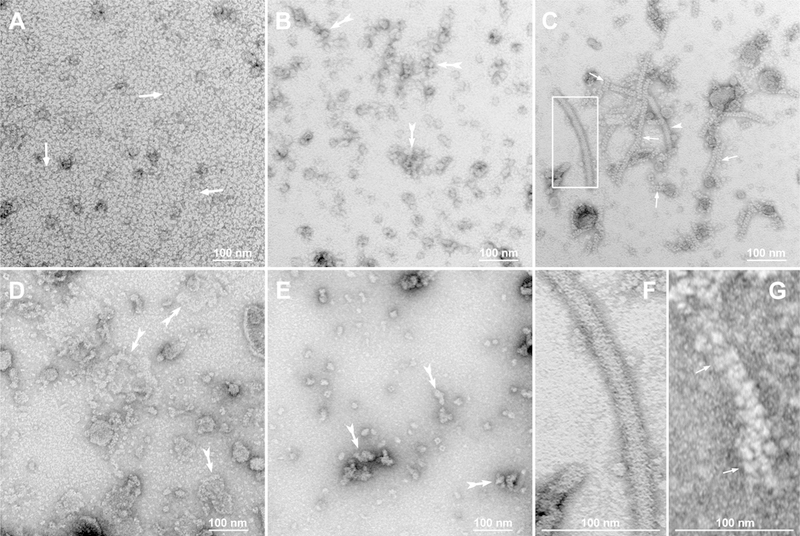
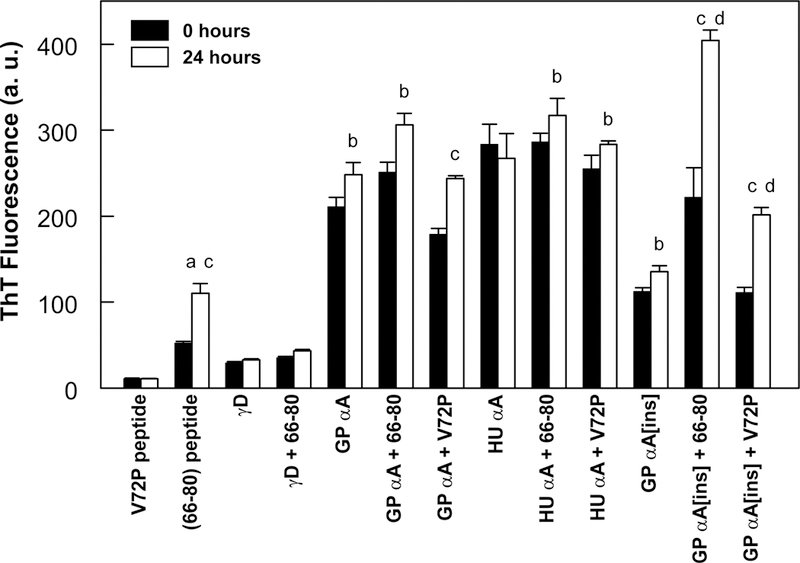
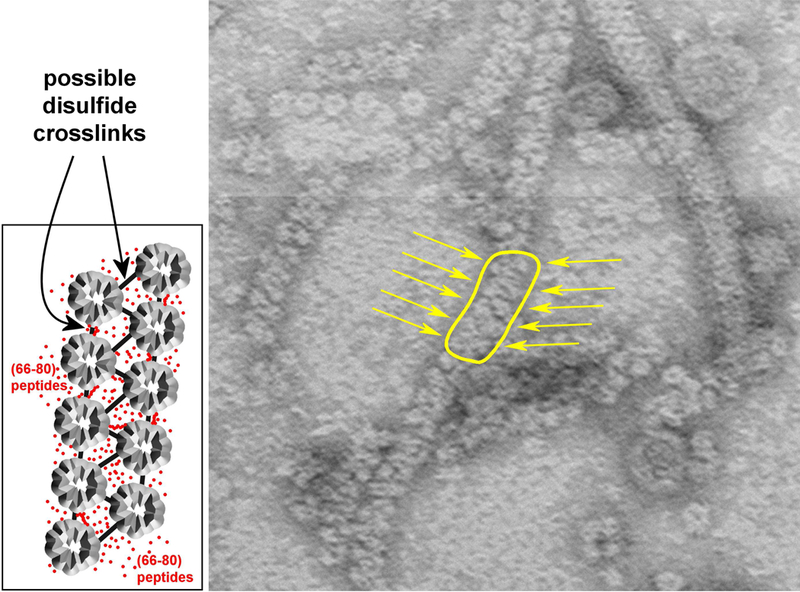
This article has been retracted: please see Elsevier Policy on Article Withdrawal (https://www.elsevier.com/about/our-business/policies/article-withdrawal). This article has been retracted at the request of the authors. The senior author contacted the journal in a forthright manner, in an effort to preserve the scientific integrity of the literature, after discovering a significant error in the results reported in the article. The authors were recently made aware of a paper by Kim et al. (Nature Commun. 2019) which shows a spirosome structure (the enzyme aldehyde-alcohol dehydrogenase) present in E. coli (Fig. 5a) that is very similar to the structure the authors thought formed when synthetic alpha A crystallin (66-80) peptide was incubated for 24 h with recombinant guinea pig alpha A insert crystallin (see Kumarasamy et al., Figs. 7C and F, and Fig. 9). Subsequent to publication of their report, the authors later found a number of images that showed what appeared to be the same structure present in samples of their presumably purified recombinant guinea pig alpha A insert crystallin which had been incubated without peptide for 24 h. Hence, the authors now conclude that the structures shown in Figs. 7C and F, and Fig. 9 of their article published in this journal are actually due to E. coli contaminant aldehyde-alcohol dehydrogenase. The authors deeply regret this error and any inconvenience it may have caused.
Keywords: Amyloid fibrils; Disulfide crosslinking; Guinea pig; Human; Lens crystallins; Nuclear cataract; Transmission electron microscopy; αA(66−80) peptide.
Copyright © 2018 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Competing Interests
The authors declare no competing interests.
Figures


Similar articles
-
Retraction notice to "Peptide-induced formation of protein aggregates and amyloid fibrils in human and Guinea pig αA-crystallins under physiological conditions of temperature and pH" [Exp. Eye Res. 179 (2019) 193-205].Exp Eye Res. 2020 May;194:107990. doi: 10.1016/j.exer.2020.107990. Epub 2020 Apr 16. Exp Eye Res. 2020. PMID: 32307097 Free PMC article.
-
Role of αA-crystallin-derived αA66-80 peptide in guinea pig lens crystallin aggregation and insolubilization.Exp Eye Res. 2015 Mar;132:151-60. doi: 10.1016/j.exer.2015.01.024. Epub 2015 Jan 29. Exp Eye Res. 2015. PMID: 25639202 Free PMC article.
-
Effect of Green Tea Polyphenol Epigallocatechin-3-gallate on the Aggregation of αA(66-80) Peptide, a Major Fragment of αA-crystallin Involved in Cataract Development.Curr Eye Res. 2017 Oct;42(10):1368-1377. doi: 10.1080/02713683.2017.1324628. Epub 2017 Jun 19. Curr Eye Res. 2017. PMID: 28628342
-
Proteostasis and the Regulation of Intra- and Extracellular Protein Aggregation by ATP-Independent Molecular Chaperones: Lens α-Crystallins and Milk Caseins.Acc Chem Res. 2018 Mar 20;51(3):745-752. doi: 10.1021/acs.accounts.7b00250. Epub 2018 Feb 14. Acc Chem Res. 2018. PMID: 29442498 Review.
-
Structure and function of α-crystallins: Traversing from in vitro to in vivo.Biochim Biophys Acta. 2016 Jan;1860(1 Pt B):149-66. doi: 10.1016/j.bbagen.2015.06.008. Epub 2015 Jun 25. Biochim Biophys Acta. 2016. PMID: 26116912 Review.
References
-
- Alliegro MC, 2000. Effects of dithiothreitol on protein activity unrelated to thiol-disulfide exchange: for consideration in the analysis of protein function with Cleland’s reagent. Analytical biochemistry 282, 102–106. - PubMed
-
- Attanasio F, Cataldo S, Fisichella S, Nicoletti S, Nicoletti VG, Pignataro B, Savarino A, Rizzarelli E, 2009. Protective effects of L- and D-carnosine on alpha-crystallin amyloid fibril formation: implications for cataract disease. Biochemistry 48, 6522–6531. - PubMed
-
- Augusteyn RC, Koretz JF, Schurtenberger P, 1989. The effect of phosphorylation on the structure of alpha-crystallin. Biochimica et biophysica acta 999, 293–299. - PubMed
-
- Balbach JJ, Ishii Y, Antzutkin ON, Leapman RD, Rizzo NW, Dyda F, Reed J, Tycko R, 2000. Amyloid fibril formation by A beta 16–22, a seven-residue fragment of the Alzheimer’s beta-amyloid peptide, and structural characterization by solid state NMR. Biochemistry 39, 13748–13759. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
