

Comparative Analysis and Refinement of Human PSC-Derived Kidney Organoid Differentiation with Single-Cell Transcriptomics
- PMID: 30449713
- PMCID: PMC6324730
- DOI: 10.1016/j.stem.2018.10.010
Comparative Analysis and Refinement of Human PSC-Derived Kidney Organoid Differentiation with Single-Cell Transcriptomics
Abstract
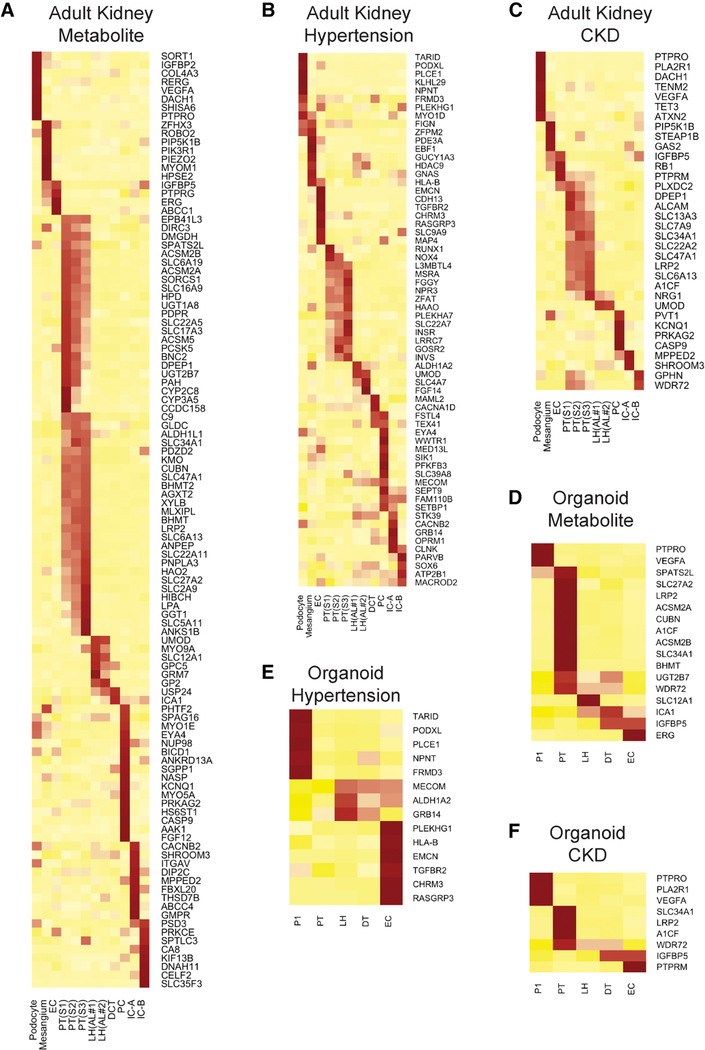
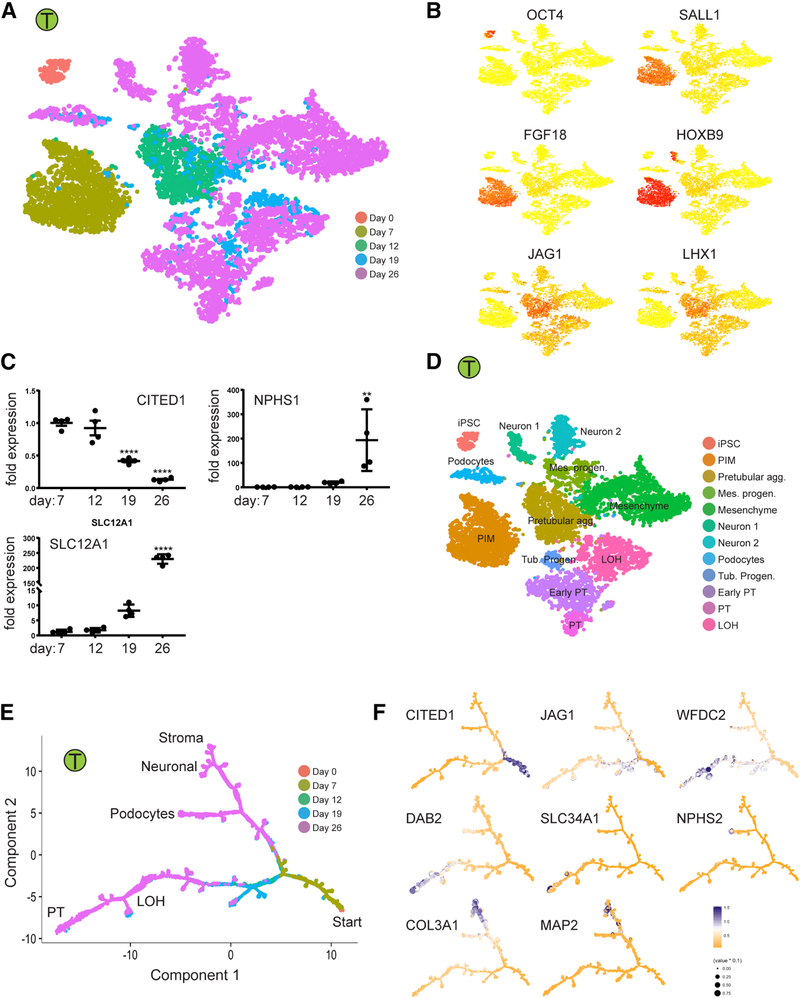
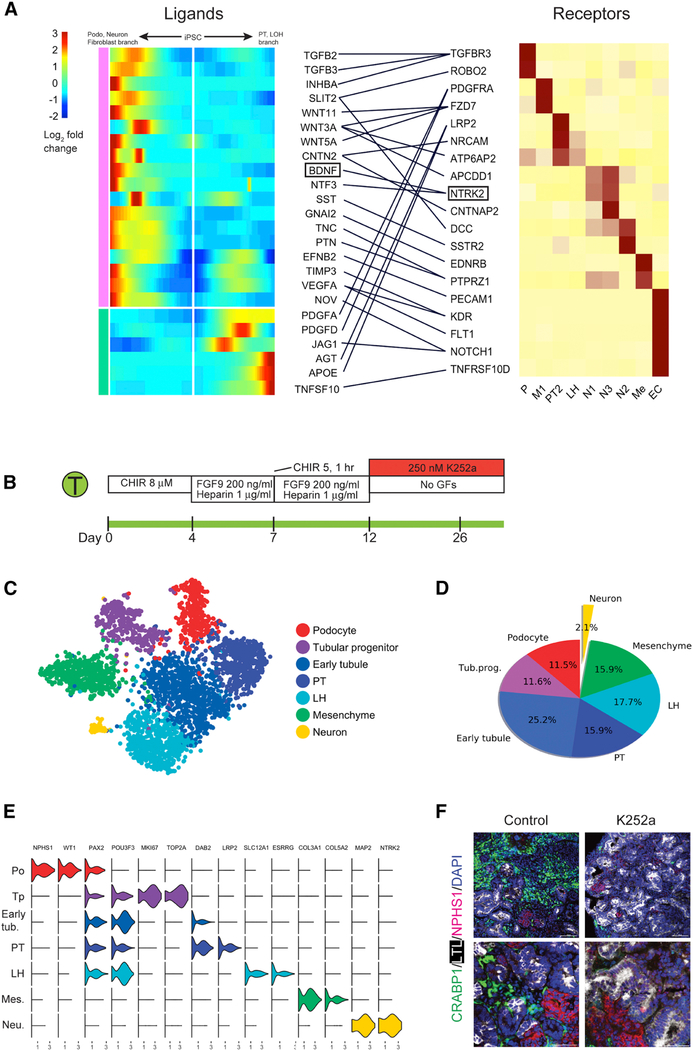
Kidney organoids derived from human pluripotent stem cells have great utility for investigating organogenesis and disease mechanisms and, potentially, as a replacement tissue source, but how closely organoids derived from current protocols replicate adult human kidney is undefined. We compared two directed differentiation protocols by single-cell transcriptomics of 83,130 cells from 65 organoids with single-cell transcriptomes of fetal and adult kidney cells. Both protocols generate a diverse range of kidney cells with differing ratios, but organoid-derived cell types are immature, and 10%-20% of cells are non-renal. Reconstructing lineage relationships by pseudotemporal ordering identified ligands, receptors, and transcription factor networks associated with fate decisions. Brain-derived neurotrophic factor (BDNF) and its cognate receptor NTRK2 were expressed in the neuronal lineage during organoid differentiation. Inhibiting this pathway improved organoid formation by reducing neurons by 90% without affecting kidney differentiation, highlighting the power of single-cell technologies to characterize and improve organoid differentiation.
Keywords: BDNF signaling; Dropseq; human kidney; induced pluripotent stem cells; kidney organoid; off-target cells; single cell RNA-seq; stem cell differentiation.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests
The authors declare no competing interests.
Figures
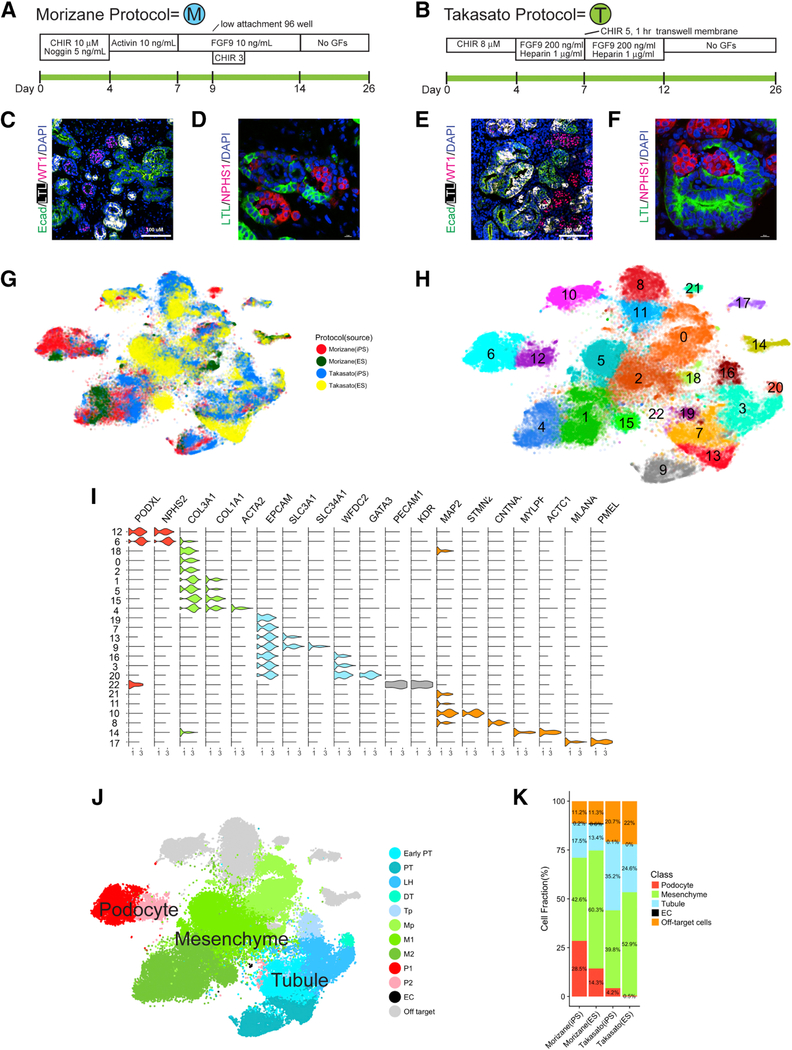
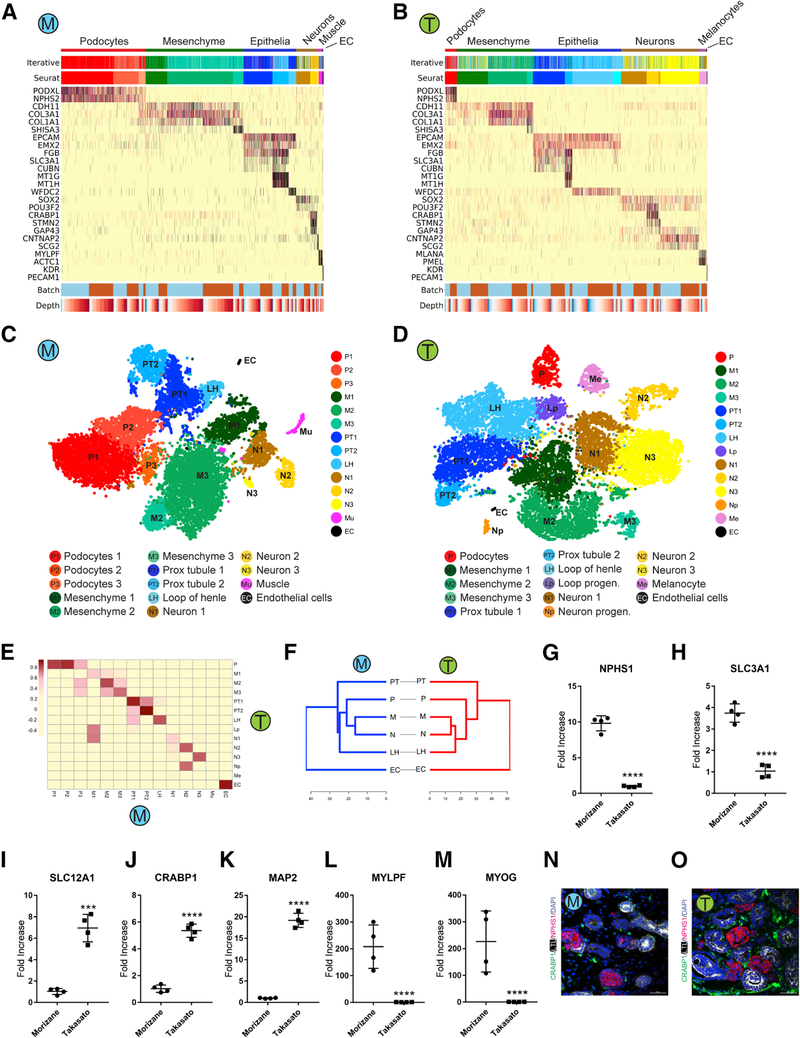
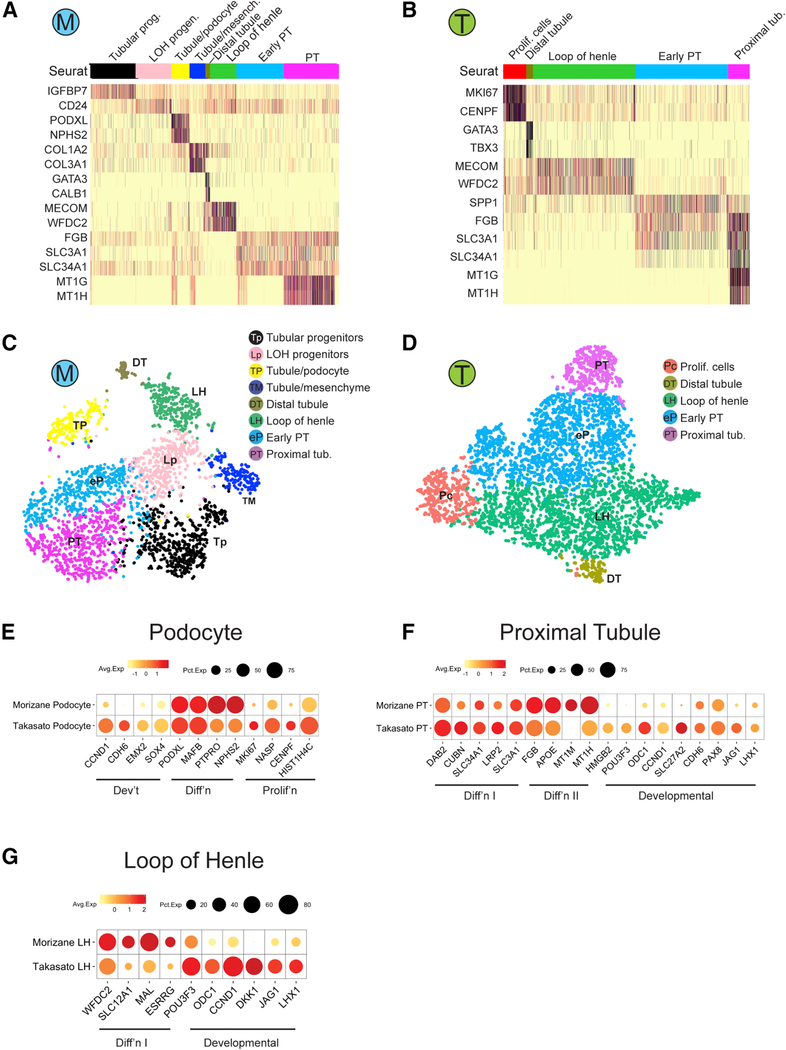
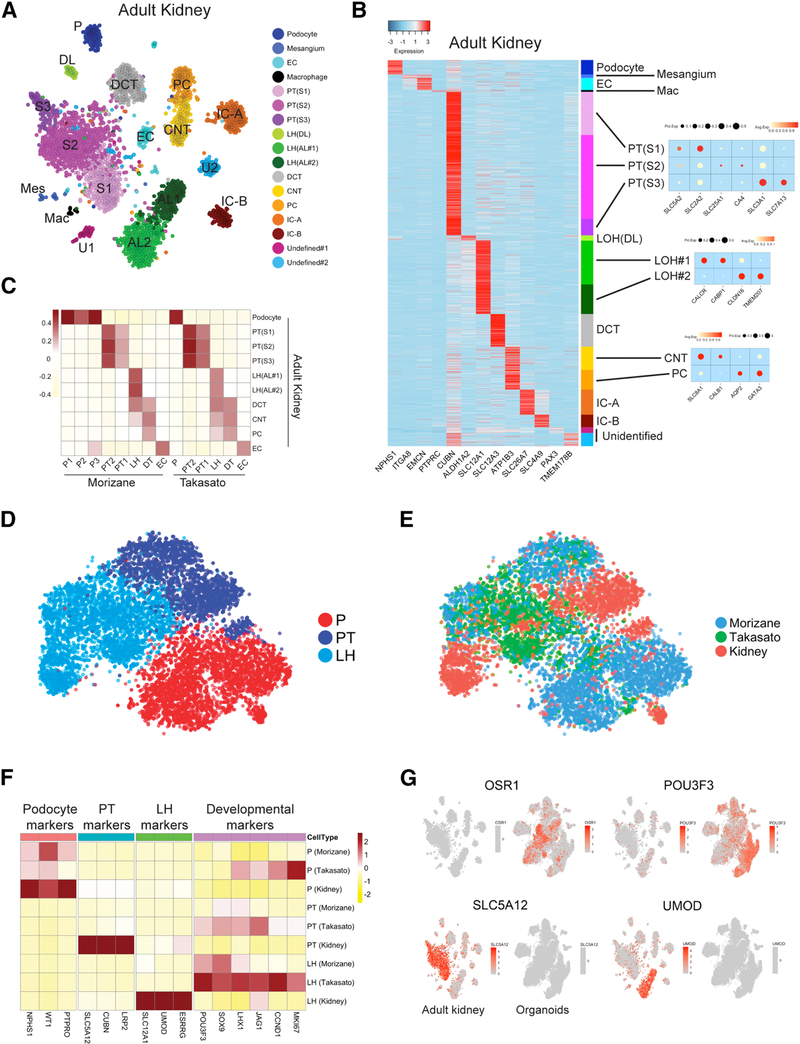
Comment in
-
Evaluation of Wu et al.: Strengthening Single-Cell Bioinformatic Comparisons for Improving Organoid Differentiation.Cell Stem Cell. 2019 Mar 7;24(3):343-344. doi: 10.1016/j.stem.2019.02.010. Cell Stem Cell. 2019. PMID: 30849359
References
-
- Bindels RJ, Hartog A, Timmermans JA, and van Os CH (1991). Immunocytochemical localization of calbindin-D28k, calbindin-D9k and parvalbumin in rat kidney. Contrib Nephrol 91, 7–13. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
