

Inhibiting the Evolution of Antibiotic Resistance
- PMID: 30449724
- PMCID: PMC6320318
- DOI: 10.1016/j.molcel.2018.10.015
Inhibiting the Evolution of Antibiotic Resistance
Abstract
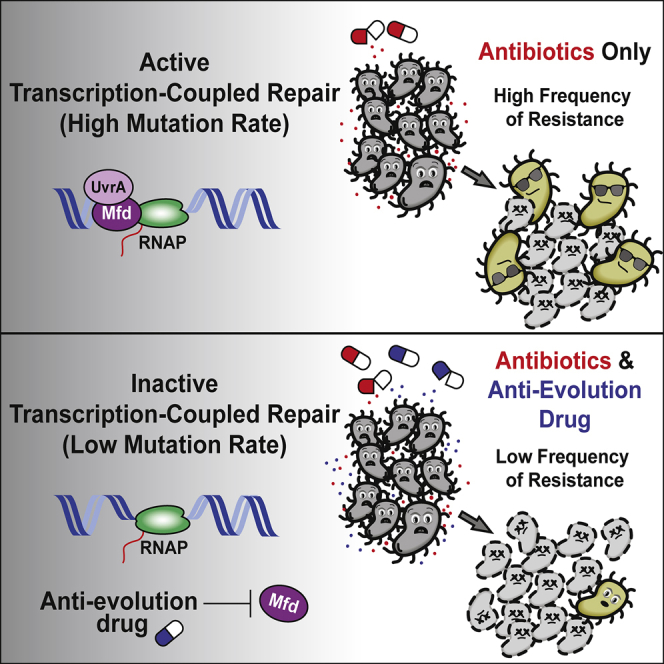
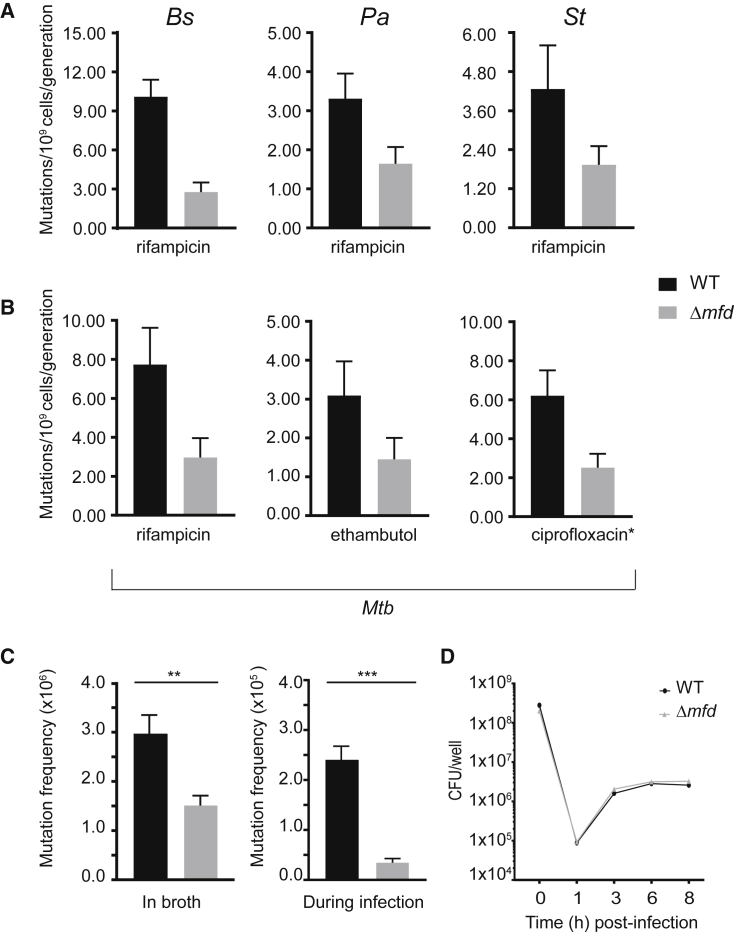
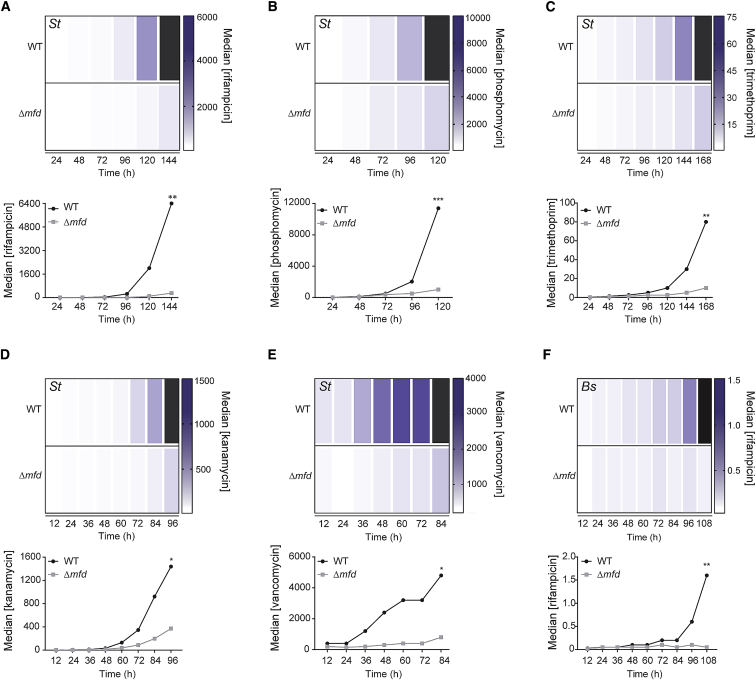
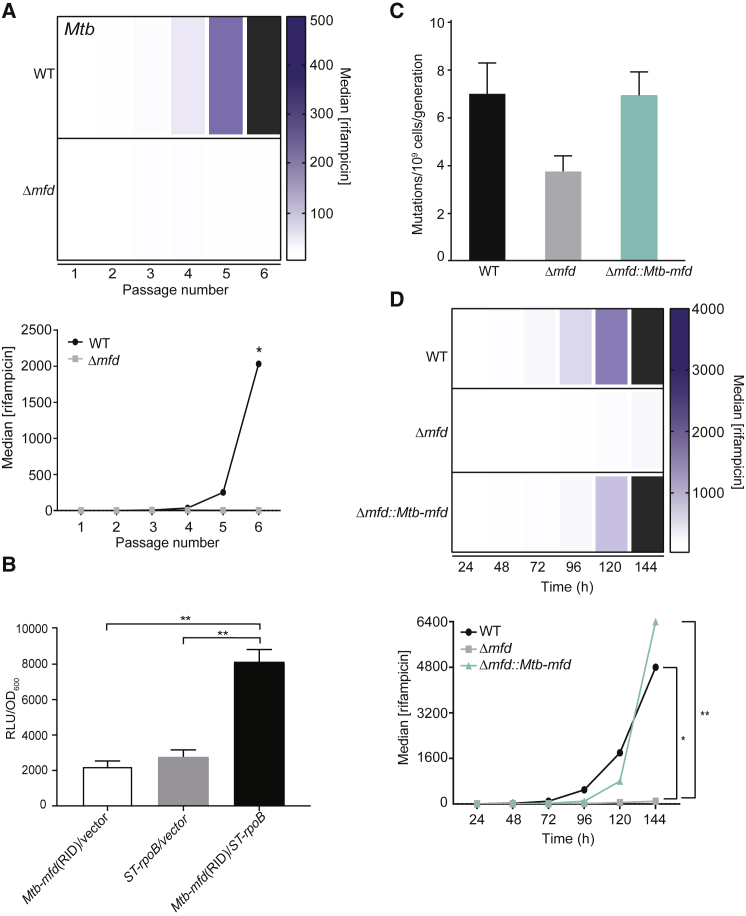
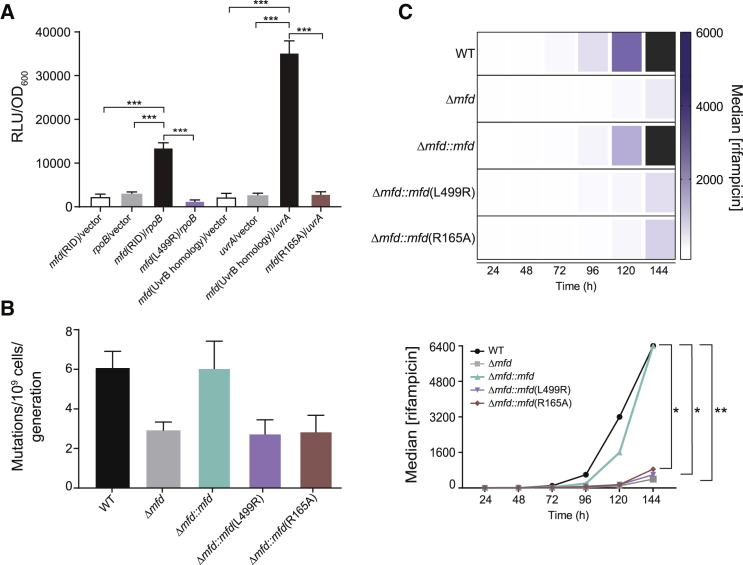
Efforts to battle antimicrobial resistance (AMR) are generally focused on developing novel antibiotics. However, history shows that resistance arises regardless of the nature or potency of new drugs. Here, we propose and provide evidence for an alternate strategy to resolve this problem: inhibiting evolution. We determined that the DNA translocase Mfd is an "evolvability factor" that promotes mutagenesis and is required for rapid resistance development to all antibiotics tested across highly divergent bacterial species. Importantly, hypermutator alleles that accelerate AMR development did not arise without Mfd, at least during evolution of trimethoprim resistance. We also show that Mfd's role in AMR development depends on its interactions with the RNA polymerase subunit RpoB and the nucleotide excision repair protein UvrA. Our findings suggest that AMR development can be inhibited through inactivation of evolvability factors (potentially with "anti-evolution" drugs)-in particular, Mfd-providing an unexplored route toward battling the AMR crisis.
Keywords: Mfd; Mycobacteria; anti-evolution; antibiotic resistance; antimicrobial resistance; evolution; hypermutator; transcription-coupled repair.
Copyright © 2018 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
Comment in
-
Reduce your speed.Nat Rev Microbiol. 2019 Jan;17(1):1. doi: 10.1038/s41579-018-0127-8. Nat Rev Microbiol. 2019. PMID: 30470813 No abstract available.
References
-
- Almeida Da Silva P.E., Palomino J.C. Molecular basis and mechanisms of drug resistance in Mycobacterium tuberculosis: classical and new drugs. J. Antimicrob. Chemother. 2011;66:1417–1430. - PubMed
-
- Blázquez J. Hypermutation as a factor contributing to the acquisition of antimicrobial resistance. Clin. Infect. Dis. 2003;37:1201–1209. - PubMed
-
- Brandis G., Pietsch F., Alemayehu R., Hughes D. Comprehensive phenotypic characterization of rifampicin resistance mutations in Salmonella provides insight into the evolution of resistance in Mycobacterium tuberculosis. J. Antimicrob. Chemother. 2015;70:680–685. - PubMed
-
- Branum M.E., Reardon J.T., Sancar A. DNA repair excision nuclease attacks undamaged DNA. A potential source of spontaneous mutations. J. Biol. Chem. 2001;276:25421–25426. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
