

Both Type I and Type II Interferons Can Activate Antitumor M1 Macrophages When Combined With TLR Stimulation
- PMID: 30450098
- PMCID: PMC6224375
- DOI: 10.3389/fimmu.2018.02520
Both Type I and Type II Interferons Can Activate Antitumor M1 Macrophages When Combined With TLR Stimulation
Abstract
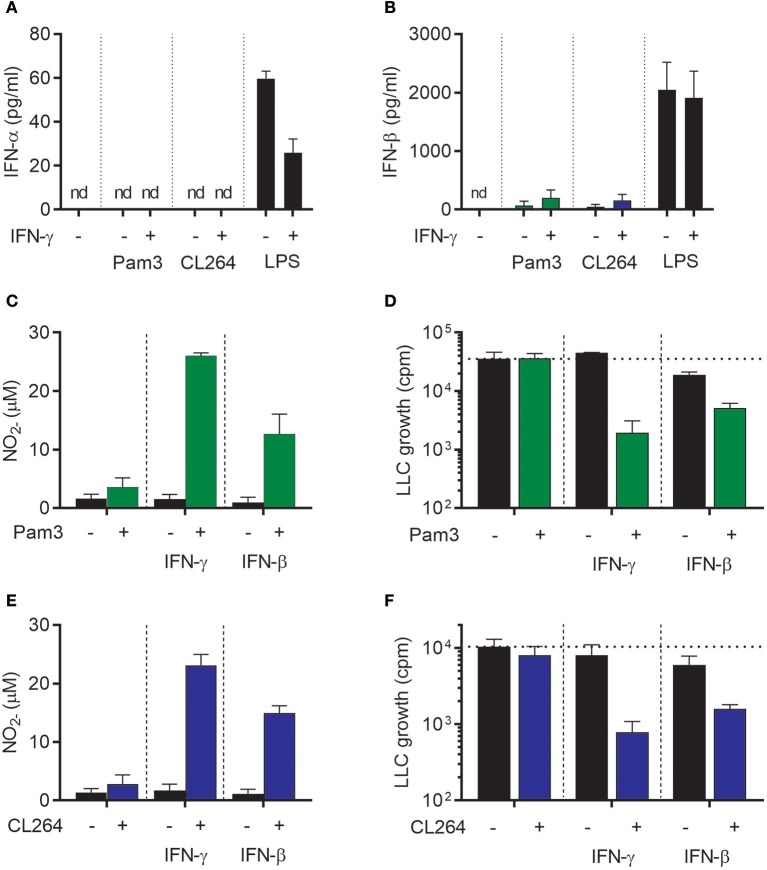
Triggering or enhancing antitumor activity of tumor-associated macrophages is an attractive strategy for cancer treatment. We have previously shown that the cytokine interferon-γ (IFN-γ), a type II IFN, could synergize with toll-like receptor (TLR) agonists for induction of antitumor M1 macrophages. However, the toxicity of IFN-γ limits its clinical use. Here, we investigated whether the less toxic type I IFNs, IFN-α, and IFN-β, could potentially replace IFN-γ for induction of antitumor M1 macrophages. We measured in vitro the ability of type I and II IFNs to synergize with TLR agonists for transcription of inducible nitric oxide synthase (iNOS) mRNA and secretion of nitric oxide (NO) by mouse bone marrow-derived macrophages (BMDMs). An in vitro growth inhibition assay was used to measure both cytotoxic and cytostatic activity of activated macrophages against Lewis lung carcinoma (LLC) cancer cells. We found that both type I and II IFNs could synergize with TLR agonists in inducing macrophage-mediated inhibition of cancer cell growth, which was dependent on NO. The ability of high dose lipopolysaccharide (LPS) to induce tumoricidal activity in macrophages in the absence of IFN-γ was shown to depend on induction of autocrine type I IFNs. Antitumor M1 macrophages could also be generated in the absence of IFN-γ by a combination of two TLR ligands when using the TLR3 agonist poly(I:C) which induces autocrine type I IFNs. Finally, we show that encapsulation of poly(I:C) into nanoparticles improved its potency to induce M1 macrophages up to 100-fold. This study reveals the potential of type I IFNs for activation of antitumor macrophages and indicates new avenues for cancer immunotherapy based on type I IFN signaling, including combination of TLR agonists.
Keywords: cancer; immunotherapy; interferon-α; interferon-β; interferon-γ; macrophages; nitric oxide; toll-like receptors.
Figures








References
-
- Bottazzi B, Polentarutti N, Acero R, Balsari A, Boraschi D, Ghezzi P, et al. Regulation of the macrophage content of neoplasms by chemoattractants. Science (1983) 220:210–2. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources