

The role of innate immunity in mucopolysaccharide diseases
- PMID: 30451296
- PMCID: PMC6590380
- DOI: 10.1111/jnc.14632
The role of innate immunity in mucopolysaccharide diseases
Abstract
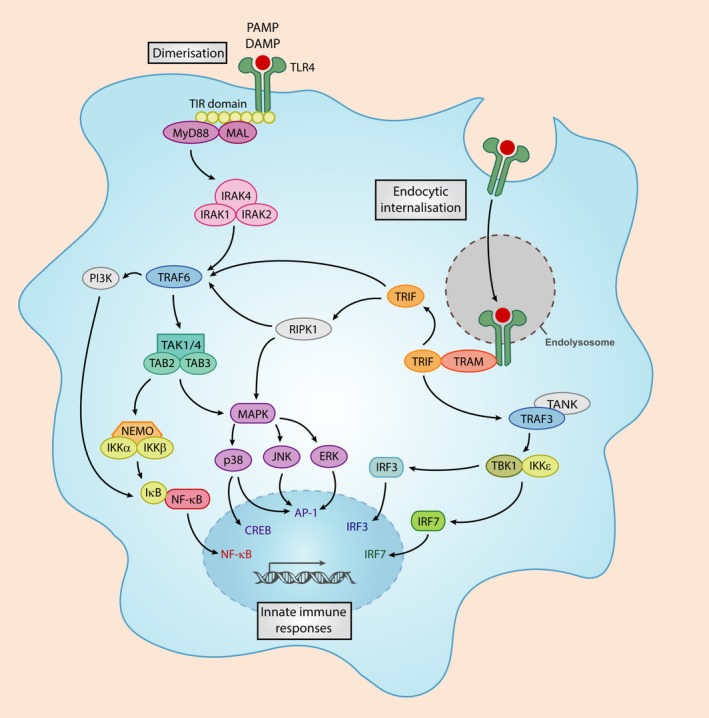
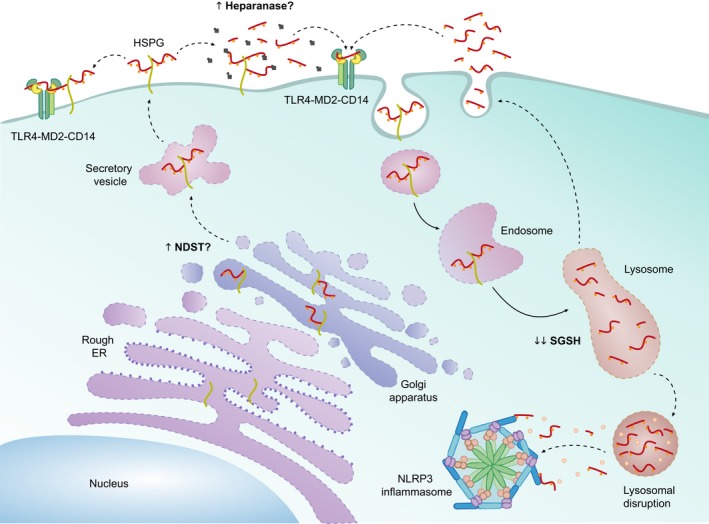
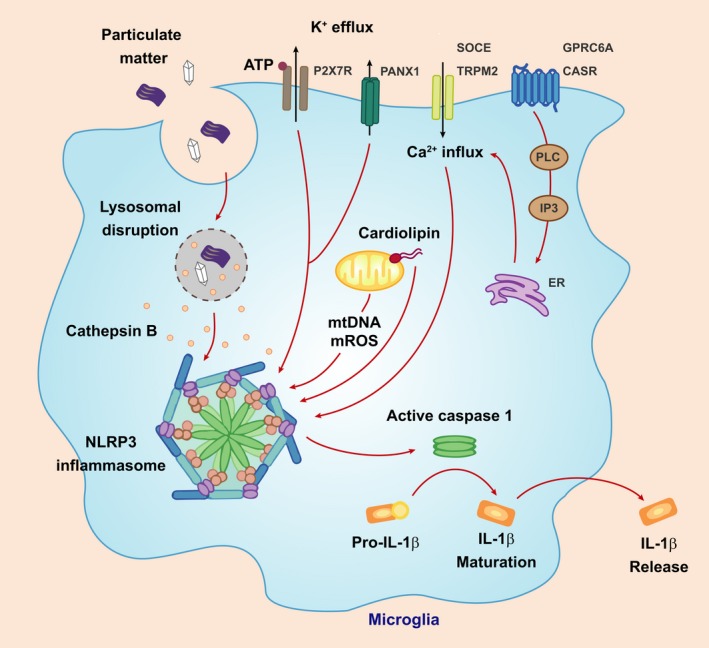
Mucopolysaccharidoses are lysosomal storage disorders characterised by accumulation of abnormal pathological glycosaminoglycans, cellular dysfunction and widespread inflammation, resulting in progressive cognitive and motor decline. Lysosomes are important mediators of immune cell function, and therefore accumulation of glycosaminoglycans (GAGs) and other abnormal substrates could affect immune function and directly impact on disease pathogenesis. This review summarises current knowledge with regard to inflammation in mucopolysaccharidosis, with an emphasis on the brain and outlines a potential role for GAGs in induction of inflammation. We propose a model by which the accumulation of GAGs and other factors may impact on innate immune signalling with particular focus on the Toll-like receptor 4 pathway. Innate immunity appears to have a dominating role in mucopolysaccharidosis; however, furthering understanding of innate immune signalling would have significant impact on highlighting novel anti-inflammatory therapeutics for use in mucopolysaccharide diseases. This article is part of the Special Issue "Lysosomal Storage Disorders".
Keywords: heparan sulphate; inflammasome; inflammation; innate immunity; lysosomal dysfunction; mucopolysaccharidosis.
© 2018 The Authors. Journal of Neurochemistry published by John Wiley & Sons Ltd on behalf of International Society for Neurochemistry.
Conflict of interest statement
BB has shares and licensed programmes in enzyme replacement stem cell gene therapy for MPSIIIA and MPSIIIB to Orchard Therapeutics Ltd. BB has shares and licensed programmes in enzyme replacement gene therapy for MPSIIIC to Phoenix Nest Inc. Neither interest competes with the content of this paper, which deals with pathophysiology and the involvement of inflammation in MPSIIIA and other LSDs. HP was supported by a PhD studentship awarded by the NRI, University of Manchester.
Figures
References
-
- Archer L. D., Langford‐Smith K. J., Bigger B. W. and Fildes J. E. (2014) Mucopolysaccharide diseases: a complex interplay between neuroinflammation, microglial activation and adaptive immunity. J. Inherit. Metab. Dis. 37, 1–12. - PubMed
-
- Arfi A., Richard M., Gandolphe C., Bonnefont‐Rousselot D., Therond P. and Scherman D. (2011) Neuroinflammatory and oxidative stress phenomena in MPS IIIA mouse model: the positive effect of long‐term aspirin treatment. Mol. Genet. Metab. 103, 18–25. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
