

The Hsp70 co-chaperone Ydj1/HDJ2 regulates ribonucleotide reductase activity
- PMID: 30452489
- PMCID: PMC6277125
- DOI: 10.1371/journal.pgen.1007462
The Hsp70 co-chaperone Ydj1/HDJ2 regulates ribonucleotide reductase activity
Abstract
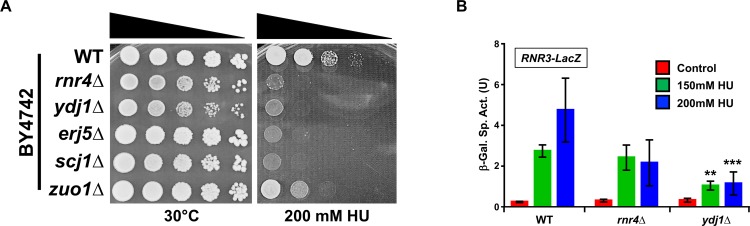
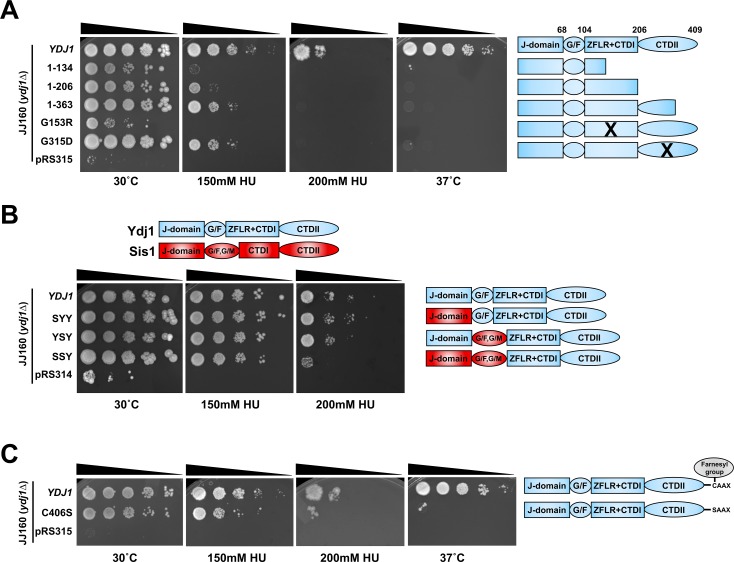
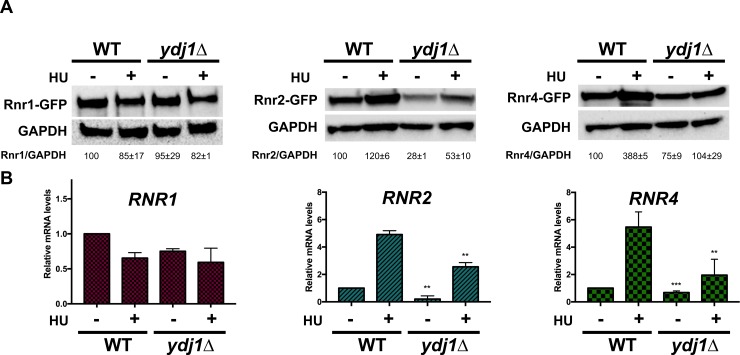
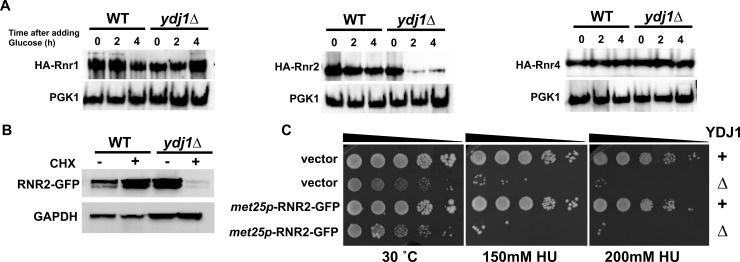
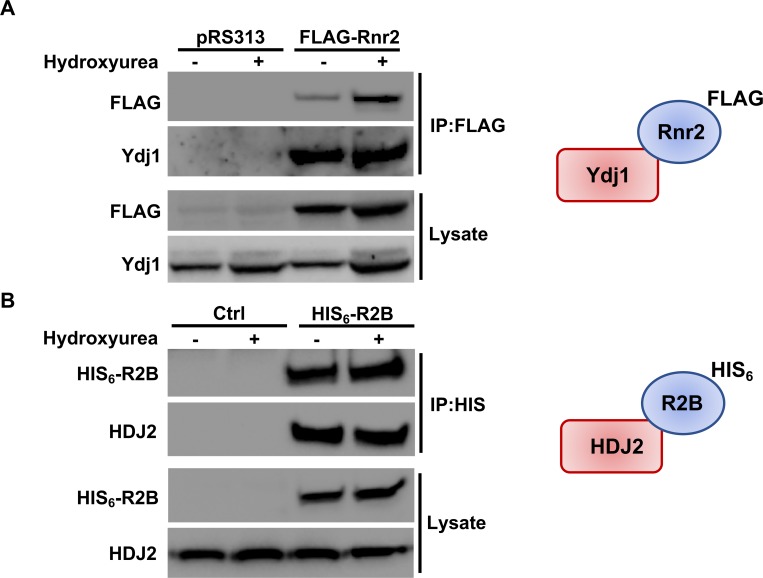
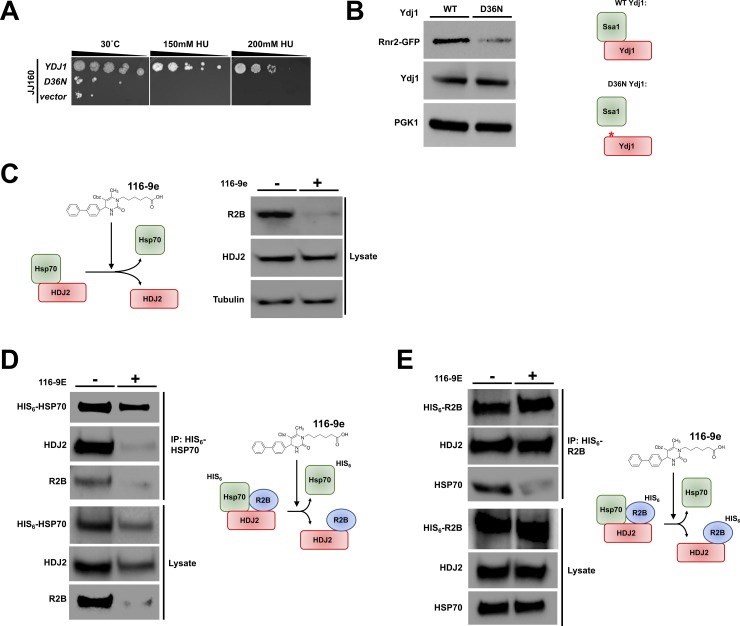
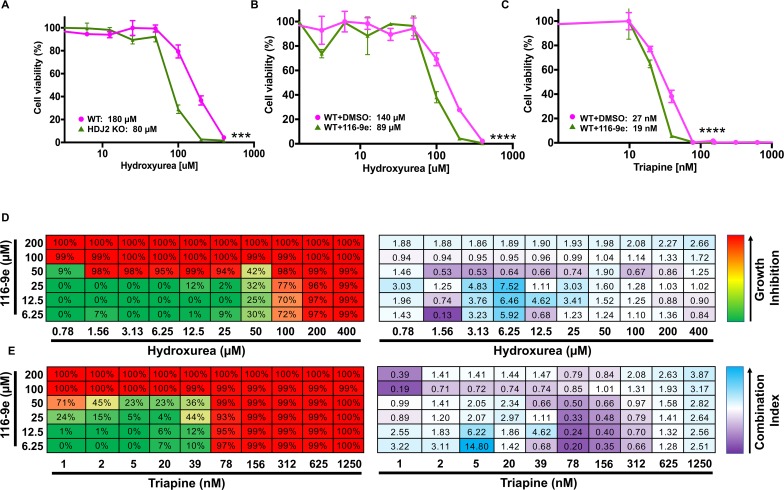
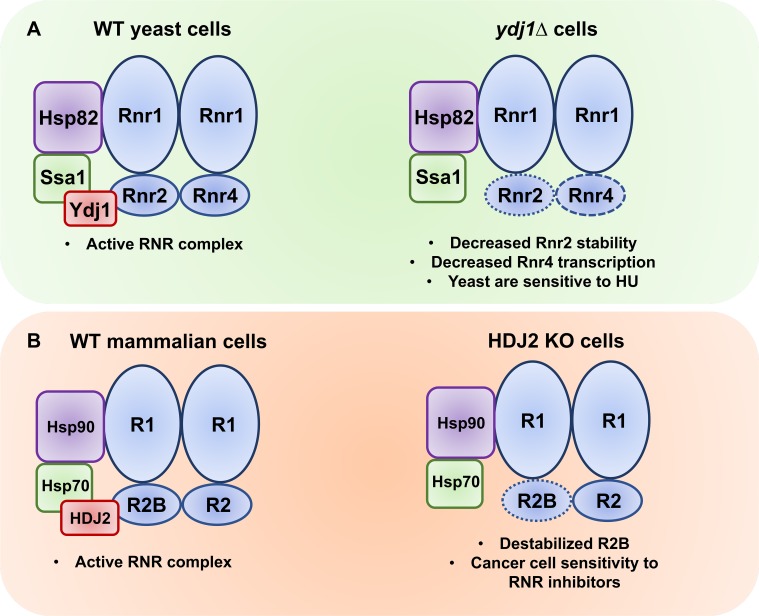
Hsp70 is a well-conserved molecular chaperone involved in the folding, stabilization, and eventual degradation of many "client" proteins. Hsp70 is regulated by a suite of co-chaperone molecules that assist in Hsp70-client interaction and stimulate the intrinsic ATPase activity of Hsp70. While previous studies have shown the anticancer target ribonucleotide reductase (RNR) is a client of Hsp70, the regulatory co-chaperones involved remain to be determined. To identify co-chaperone(s) involved in RNR activity, 28 yeast co-chaperone knockout mutants were screened for sensitivity to the RNR-perturbing agent Hydroxyurea. Ydj1, an important cytoplasmic Hsp70 co-chaperone was identified to be required for growth on HU. Ydj1 bound the RNR subunit Rnr2 and cells lacking Ydj1 showed a destabilized RNR complex. Suggesting broad conservation from yeast to human, HDJ2 binds R2B and regulates RNR stability in human cells. Perturbation of the Ssa1-Ydj1 interaction through mutation or Hsp70-HDJ2 via the small molecule 116-9e compromised RNR function, suggesting chaperone dependence of this novel role. Mammalian cells lacking HDJ2 were significantly more sensitive to RNR inhibiting drugs such as hydroxyurea, gemcitabine and triapine. Taken together, this work suggests a novel anticancer strategy-inhibition of RNR by targeting Hsp70 co-chaperone function.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Craig EA, Marszalek J. How Do J-Proteins Get Hsp70 to Do So Many Different Things? Trends Biochem Sci. 2017;42(5):355–68. Epub 2017/03/21. 10.1016/j.tibs.2017.02.007 ; PubMed Central PMCID: PMCPMC5409888. - DOI - PMC - PubMed
-
- Kim YE, Hipp MS, Bracher A, Hayer-Hartl M, Hartl FU. Molecular chaperone functions in protein folding and proteostasis. Annual review of biochemistry. 2013;82:323–55. Epub 2013/06/12. 10.1146/annurev-biochem-060208-092442 . - DOI - PubMed
-
- Nillegoda NB, Wentink AS, Bukau B. Protein Disaggregation in Multicellular Organisms. Trends Biochem Sci. 2018;43(4):285–300. Epub 2018/03/05. 10.1016/j.tibs.2018.02.003 . - DOI - PubMed
-
- Lianos GD, Alexiou GA, Mangano A, Mangano A, Rausei S, Boni L, et al. The role of heat shock proteins in cancer. Cancer letters. 2015;360(2):114–8. Epub 2015/02/28. 10.1016/j.canlet.2015.02.026 . - DOI - PubMed
-
- Shevtsov M, Huile G, Multhoff G. Membrane heat shock protein 70: a theranostic target for cancer therapy. Philosophical transactions of the Royal Society of London Series B, Biological sciences. 2018;373(1738). Epub 2017/12/06. 10.1098/rstb.2016.0526 ; PubMed Central PMCID: PMCPMC5717526. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
