

Two-carbon folate cycle of commensal Lactobacillus reuteri 6475 gives rise to immunomodulatory ethionine, a source for histone ethylation
- PMID: 30452879
- PMCID: PMC6404577
- DOI: 10.1096/fj.201801848R
Two-carbon folate cycle of commensal Lactobacillus reuteri 6475 gives rise to immunomodulatory ethionine, a source for histone ethylation
Abstract
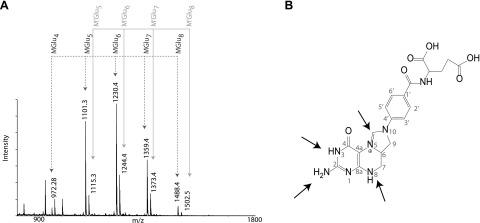
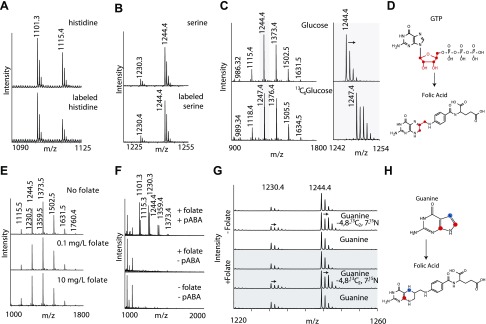
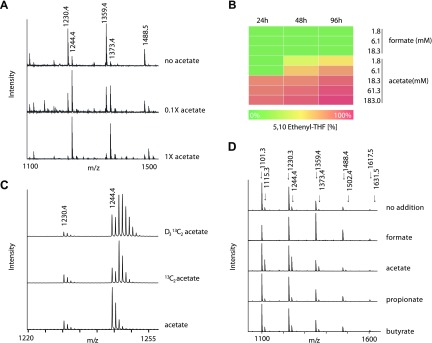
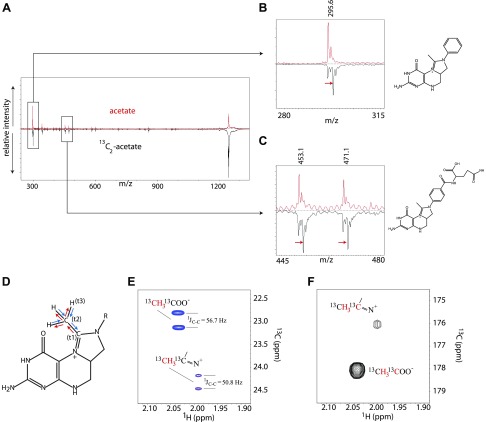
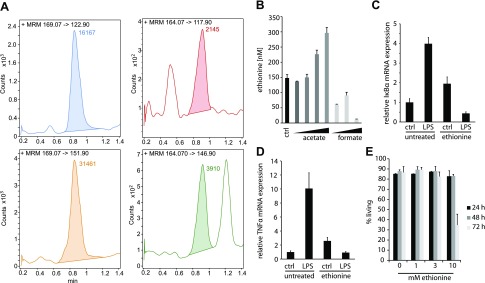
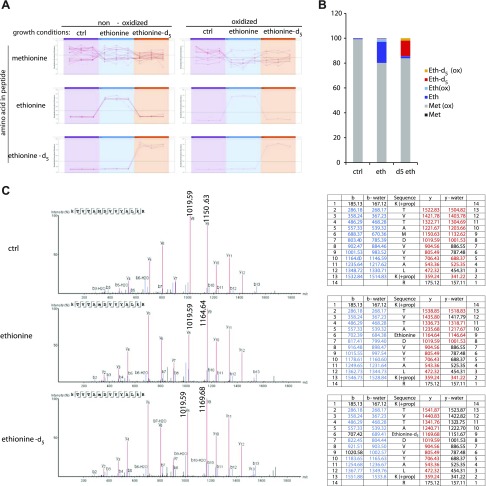
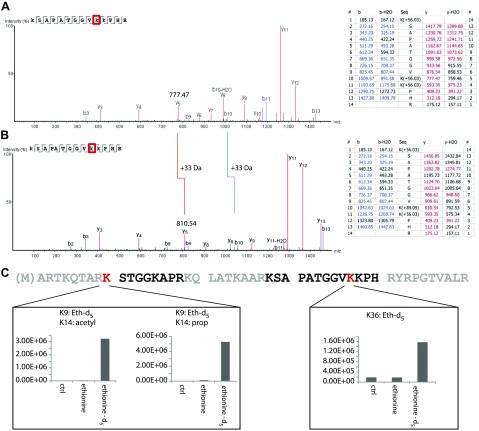
Colonization of the gut by certain probiotic Lactobacillus reuteri strains has been associated with reduced risk of inflammatory diseases and colorectal cancer. Previous studies pointed to a functional link between immunomodulation, histamine production, and folate metabolism, the central 1-carbon pathway for the transfer of methyl groups. Using mass spectrometry and NMR spectroscopy, we analyzed folate metabolites of L. reuteri strain 6475 and discovered that the bacterium produces a 2-carbon-transporting folate in the form of 5,10-ethenyl-tetrahydrofolyl polyglutamate. Isotopic labeling permitted us to trace the source of the 2-carbon unit back to acetate of the culture medium. We show that the 2C folate cycle of L. reuteri is capable of transferring 2 carbon atoms to homocysteine to generate the unconventional amino acid ethionine, a known immunomodulator. When we treated monocytic THP-1 cells with ethionine, their transcription of TNF-α was inhibited and cell proliferation reduced. Mass spectrometry of THP-1 histones revealed incorporation of ethionine instead of methionine into proteins, a reduction of histone-methylation, and ethylation of histone lysine residues. Our findings suggest that the microbiome can expose the host to ethionine through a novel 2-carbon transporting variant of the folate cycle and modify human chromatin via ethylation.-Röth, D., Chiang, A. J., Hu, W., Gugiu, G. B., Morra, C. N., Versalovic, J., Kalkum, M. The two-carbon folate cycle of commensal Lactobacillus reuteri 6475 gives rise to immunomodulatory ethionine, a source for histone ethylation.
Keywords: ethenyltetrahydrofolate; lysine ethylation; microbiome; posttranslational modification; probiotic bacteria.
Conflict of interest statement
D.R., A.J.C., C.N.M., J.V., and M.K. were supported by U.S. National Institutes of Health (NIH), National Cancer Institute Grant U01 CA170930 (to J.V.). The use of the City of Hope Mass Spectrometry and Proteomics core facility was partially supported by NIH, National Cancer Institute Grant P30 CA33572. The authors declare no conflicts of interest.
Figures
References
-
- Food and Agriculture Organization of the United Nations WHO (2006) Probiotics in Food: Health and Nutritional Properties and Guidelines for Evaluation, Food and Agriculture Organization of the United Nations WHO, Rome, Italy
-
- Saarela M., Lähteenmäki L., Crittenden R., Salminen S., Mattila-Sandholm T. (2002) Gut bacteria and health foods--the European perspective. Int. J. Food Microbiol. 78, 99–117 - PubMed
-
- Walter J., Chagnaud P., Tannock G. W., Loach D. M., Dal Bello F., Jenkinson H. F., Hammes W. P., Hertel C. (2005) A high-molecular-mass surface protein (Lsp) and methionine sulfoxide reductase B (MsrB) contribute to the ecological performance of Lactobacillus reuteri in the murine gut. Appl. Environ. Microbiol. 71, 979–986 - PMC - PubMed
-
- Reuter G. (2001) The Lactobacillus and Bifidobacterium microflora of the human intestine: composition and succession. Curr. Issues Intest. Microbiol. 2, 43–53 - PubMed
-
- Zella G. C., Hait E. J., Glavan T., Gevers D., Ward D. V., Kitts C. L., Korzenik J. R. (2011) Distinct microbiome in pouchitis compared to healthy pouches in ulcerative colitis and familial adenomatous polyposis. Inflamm. Bowel Dis. 17, 1092–1100 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
