

Exquisite ligand stereoselectivity of a Drosophila juvenile hormone receptor contrasts with its broad agonist repertoire
- PMID: 30455350
- PMCID: PMC6333893
- DOI: 10.1074/jbc.RA118.005992
Exquisite ligand stereoselectivity of a Drosophila juvenile hormone receptor contrasts with its broad agonist repertoire
Abstract
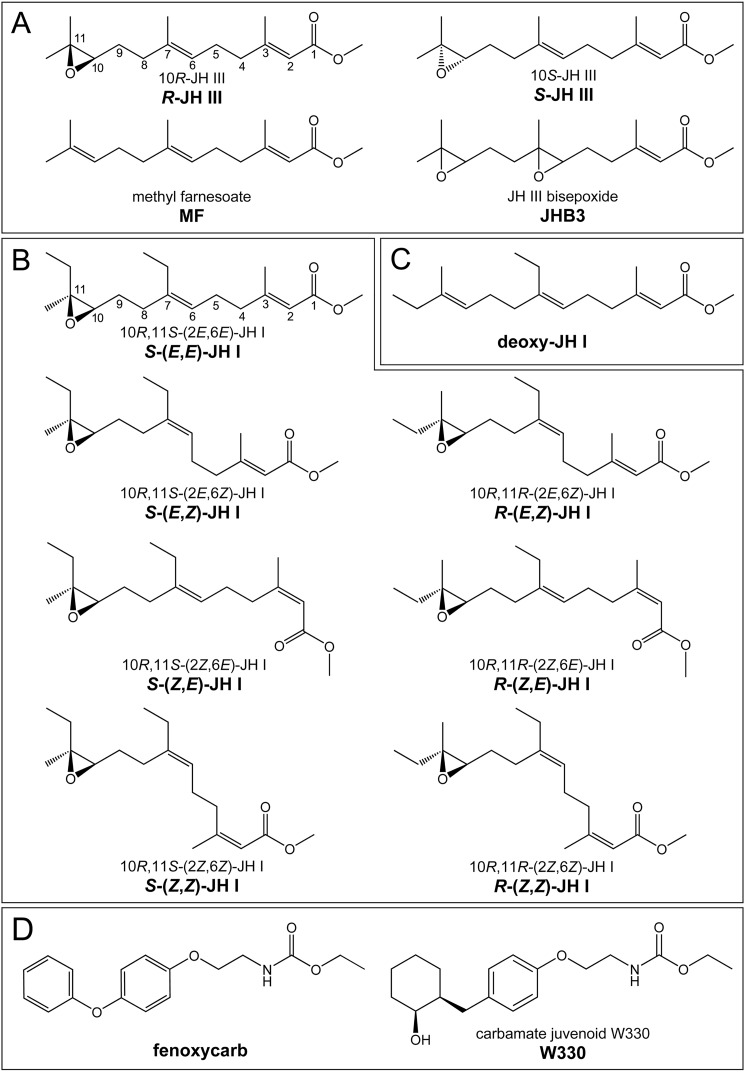
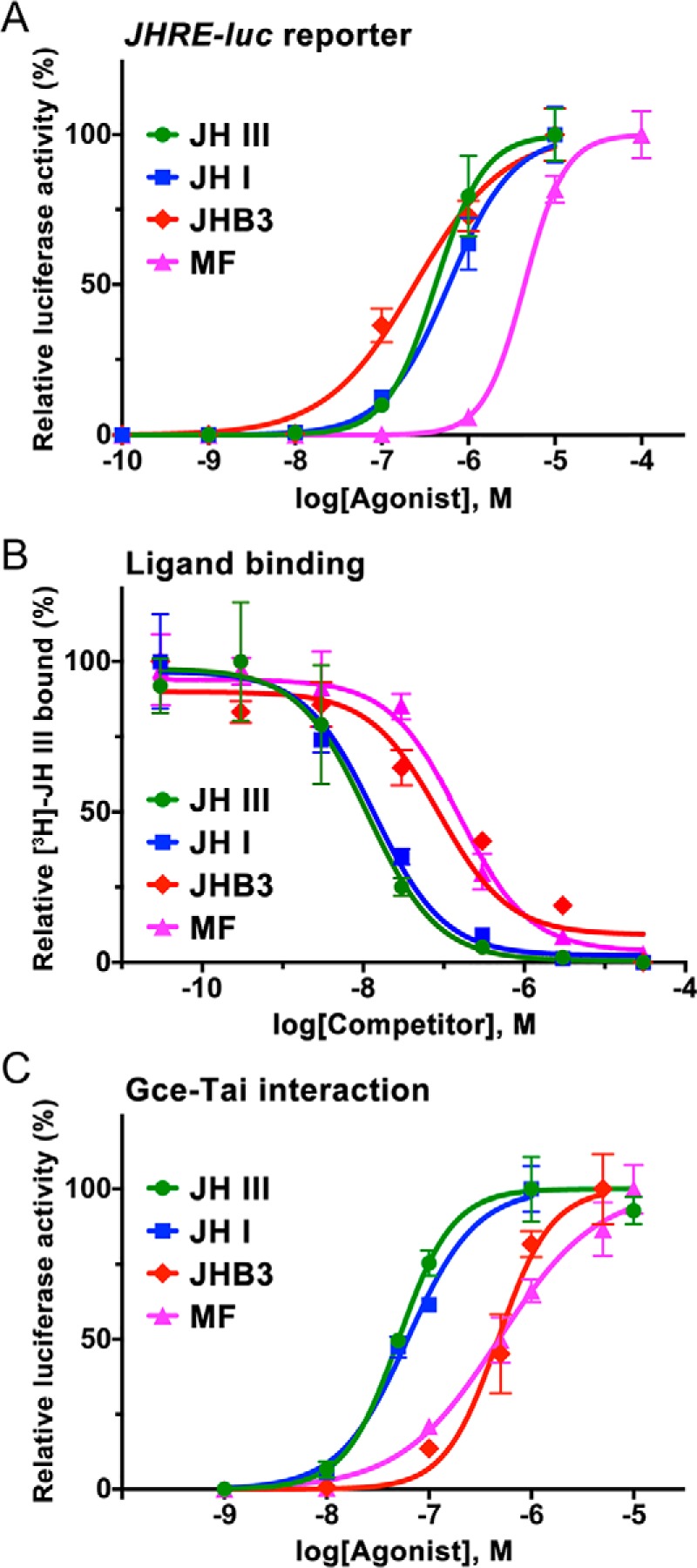
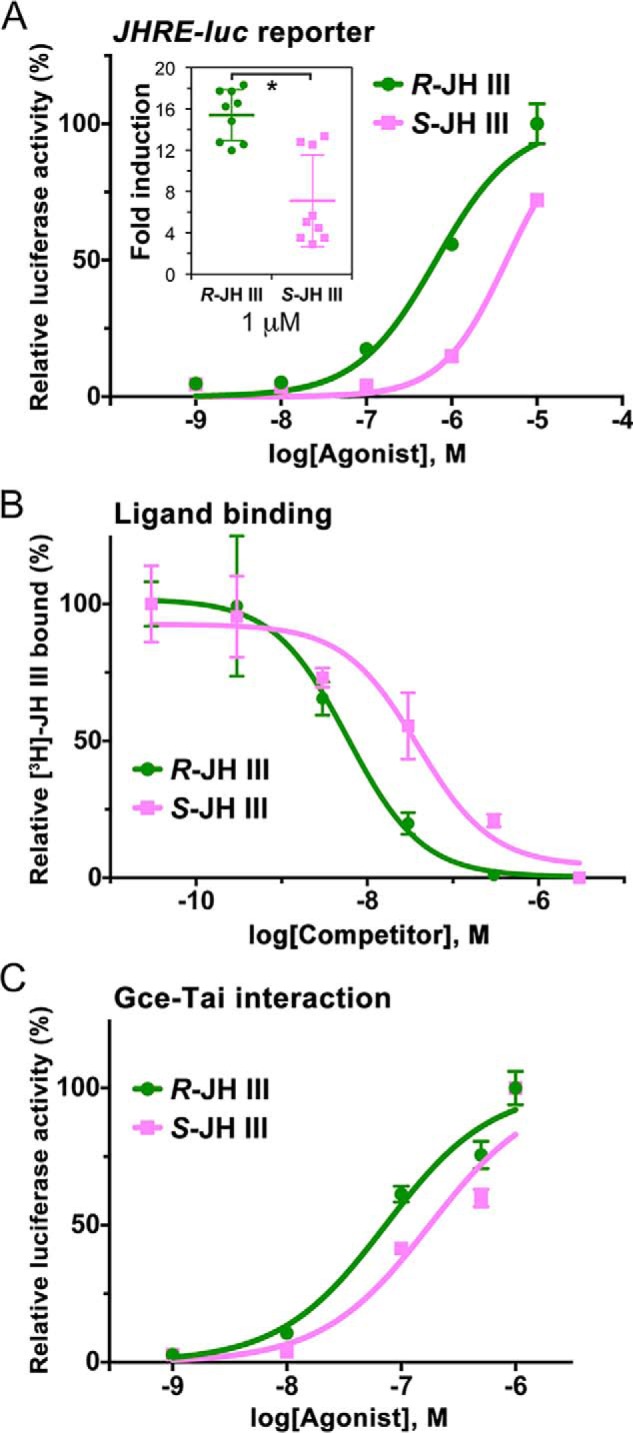
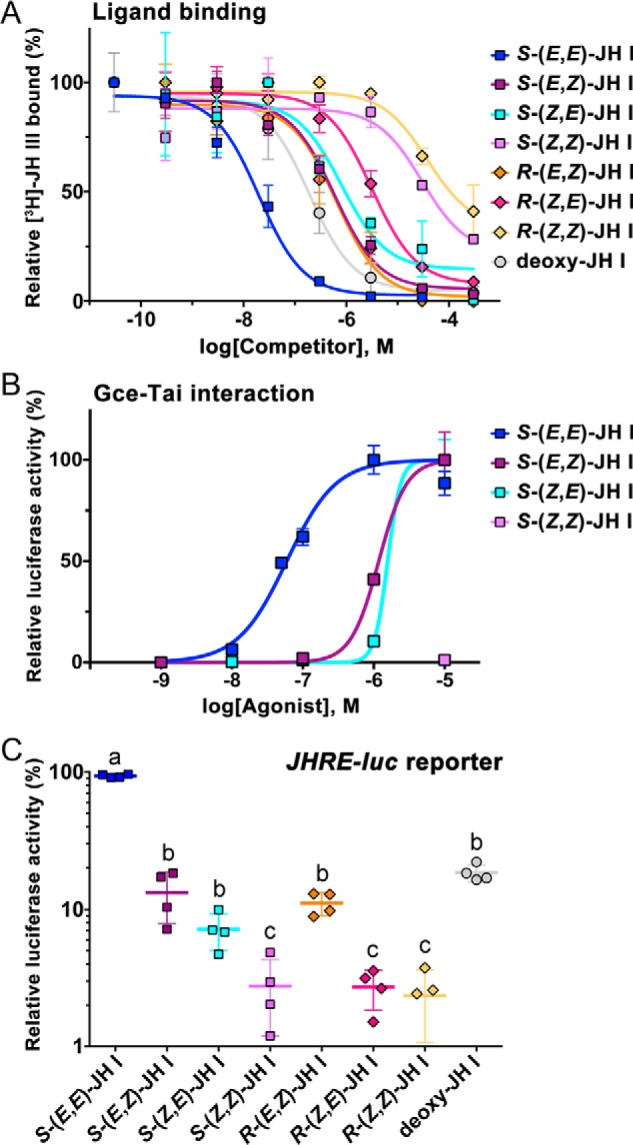
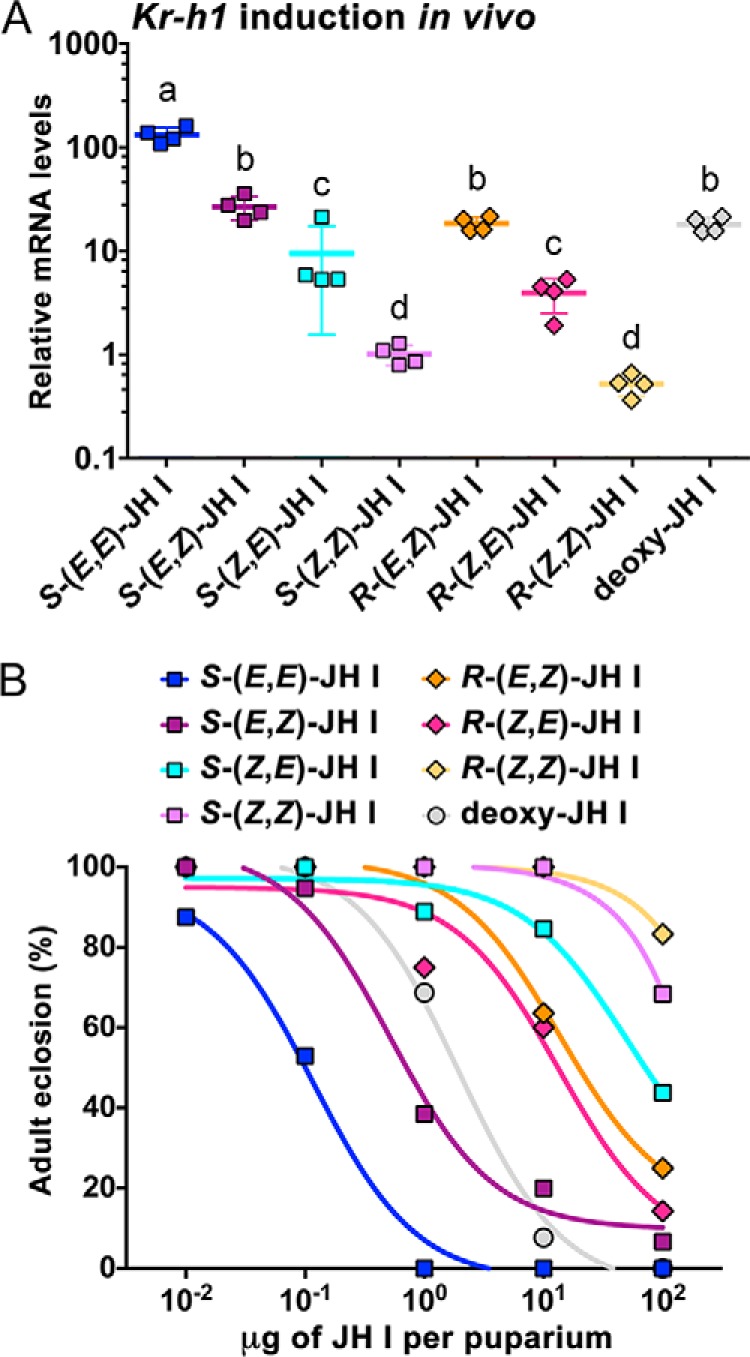
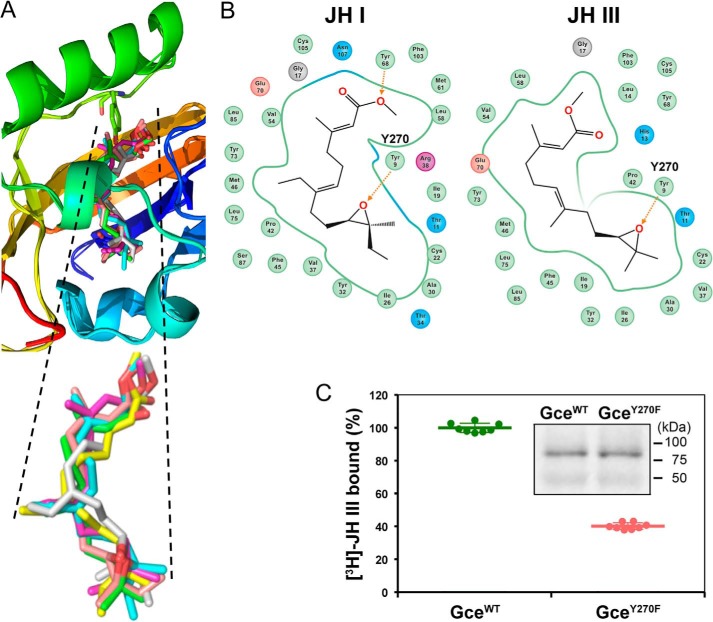
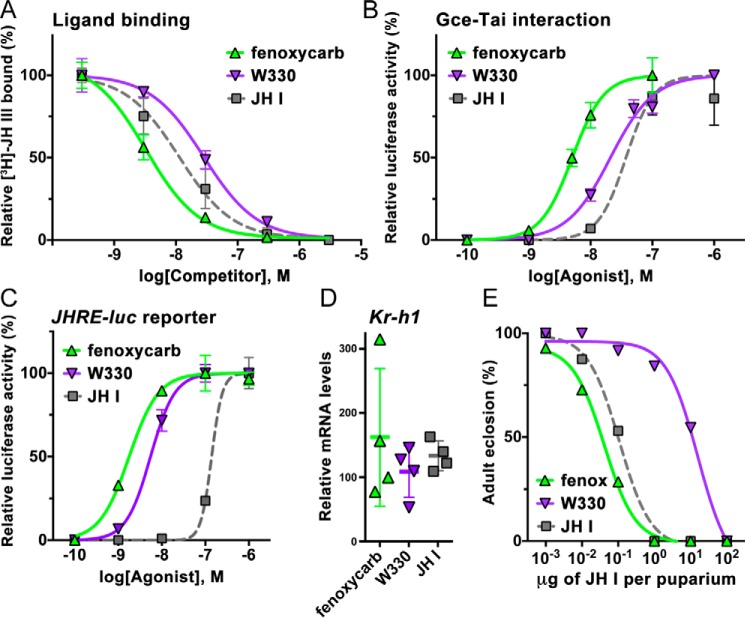
The sesquiterpenoid juvenile hormone (JH) is vital to insect development and reproduction. Intracellular JH receptors have recently been established as basic helix-loop-helix transcription factor (bHLH)/PAS proteins in Drosophila melanogaster known as germ cell-expressed (Gce) and its duplicate paralog, methoprene-tolerant (Met). Upon binding JH, Gce/Met activates its target genes. Insects possess multiple native JH homologs whose molecular activities remain unexplored, and diverse synthetic compounds including insecticides exert JH-like effects. How the JH receptor recognizes its ligands is unknown. To determine which structural features define an active JH receptor agonist, we tested several native JHs and their nonnative geometric and optical isomers for the ability to bind the Drosophila JH receptor Gce, to induce Gce-dependent transcription, and to affect the development of the fly. Our results revealed high ligand stereoselectivity of the receptor. The geometry of the JH skeleton, dictated by two stereogenic double bonds, was the most critical feature followed by the presence of an epoxide moiety at a terminal position. The optical isomerism at carbon C11 proved less important even though Gce preferentially bound a natural JH enantiomer. The results of receptor-ligand-binding and cell-based gene activation assays tightly correlated with the ability of different geometric JH isomers to induce gene expression and morphogenetic effects in the developing insects. Molecular modeling supported the requirement for the proper double-bond geometry of JH, which appears to be its major selective mechanism. The strict stereoselectivity of Gce toward the natural hormone contrasts with the high potency of synthetic Gce agonists of disparate chemistries.
Keywords: Drosophila; basic helix-loop-helix transcription factor (bHLH); development; hormone receptor; insect; juvenile hormone (JH); ligand-binding protein; reproduction; stereoselectivity.
© 2019 Bittova et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
