

Auditory Thalamostriatal and Corticostriatal Pathways Convey Complementary Information about Sound Features
- PMID: 30459227
- PMCID: PMC6325256
- DOI: 10.1523/JNEUROSCI.1188-18.2018
Auditory Thalamostriatal and Corticostriatal Pathways Convey Complementary Information about Sound Features
Abstract
Multiple parallel neural pathways link sound-related signals to behavioral responses. For instance, the striatum, a brain structure involved in action selection and reward-related learning, receives neuronal projections from both the auditory thalamus and auditory cortex. It is not clear whether sound information that reaches the striatum through these two pathways is redundant or complementary. We used an optogenetic approach in awake mice of both sexes to identify thalamostriatal and corticostriatal neurons during extracellular recordings, and characterized neural responses evoked by sounds of different frequencies and amplitude modulation rates. We found that neurons in both pathways encode sound frequency with similar fidelity, but display different coding strategies for amplitude modulated noise. Whereas corticostriatal neurons provide a more accurate representation of amplitude modulation rate in their overall firing rate, thalamostriatal neurons convey information about the precise timing of acoustic events. These results demonstrate that auditory thalamus and auditory cortex neurons provide complementary information to the striatum, and suggest that these pathways could be differentially recruited depending on the requirements of a sound-driven behavior.SIGNIFICANCE STATEMENT Sensory signals from the cerebral cortex and the thalamus converge onto the striatum, a nucleus implicated in reward-related learning. It is not clear whether these two sensory inputs convey redundant or complementary information. By characterizing the sound-evoked responses of thalamostriatal and corticostriatal neurons, our work demonstrates that these neural pathways convey complementary information about the temporal features of sounds. This work opens new avenues for investigating how these pathways could be selectively recruited depending on task demands, and provides a framework for studying convergence of cortical and thalamic information onto the striatum in other sensory systems.
Keywords: amplitude modulation; auditory cortex; auditory thalamus; neural coding; pathway-specific; striatum.
Copyright © 2019 the authors 0270-6474/19/390271-10$15.00/0.
Figures
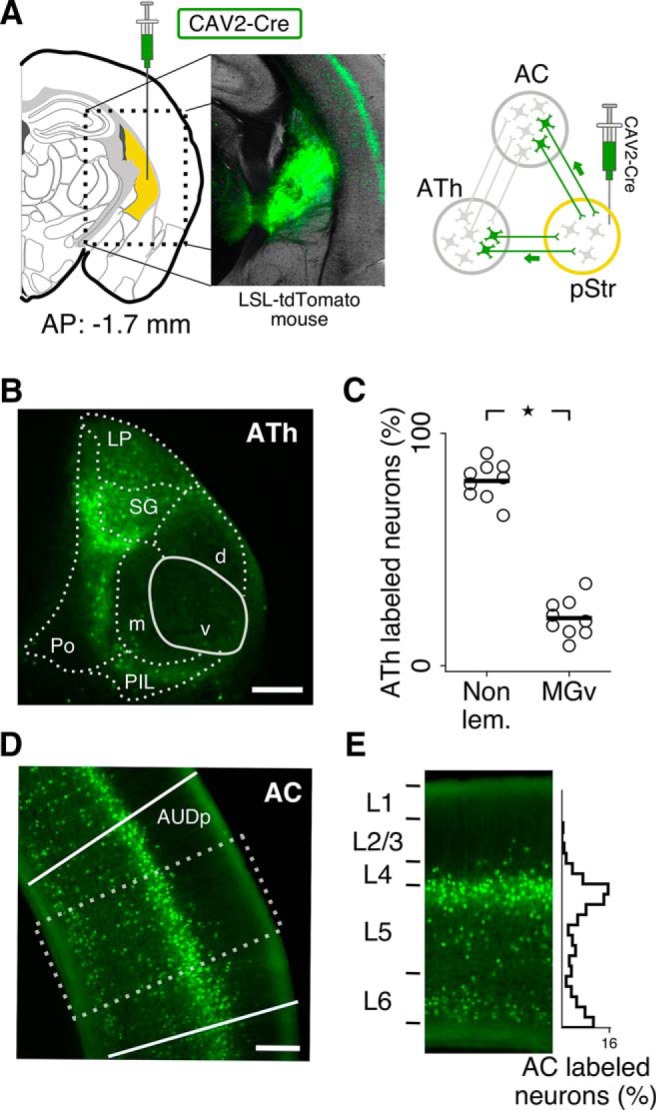
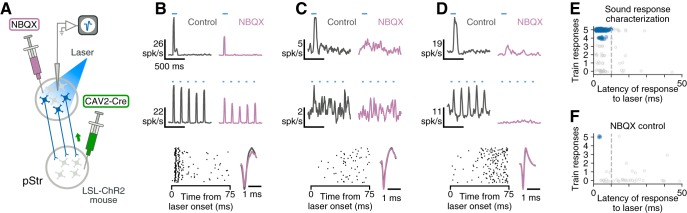
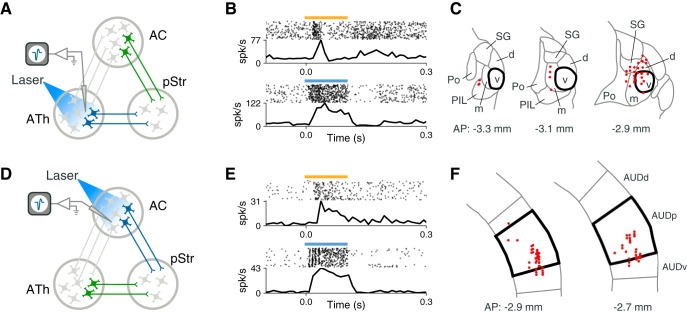


Similar articles
-
Choice-Selective Neurons in the Auditory Cortex and in Its Striatal Target Encode Reward Expectation.J Neurosci. 2019 May 8;39(19):3687-3697. doi: 10.1523/JNEUROSCI.2585-18.2019. Epub 2019 Mar 5. J Neurosci. 2019. PMID: 30837264 Free PMC article.
-
Medial geniculate body and primary auditory cortex differentially contribute to striatal sound representations.Nat Commun. 2019 Jan 24;10(1):418. doi: 10.1038/s41467-019-08350-7. Nat Commun. 2019. PMID: 30679433 Free PMC article.
-
Input-Specific Gain Modulation by Local Sensory Context Shapes Cortical and Thalamic Responses to Complex Sounds.Neuron. 2016 Jul 20;91(2):467-81. doi: 10.1016/j.neuron.2016.05.041. Epub 2016 Jun 23. Neuron. 2016. PMID: 27346532 Free PMC article.
-
Neural coding of temporal information in auditory thalamus and cortex.Neuroscience. 2008 Nov 19;157(2):484-94. doi: 10.1016/j.neuroscience.2008.07.050. Neuroscience. 2008. PMID: 19143093 Review.
-
Neural coding of temporal information in auditory thalamus and cortex.Neuroscience. 2008 Jun 12;154(1):294-303. doi: 10.1016/j.neuroscience.2008.03.065. Epub 2008 Apr 7. Neuroscience. 2008. Corrected and republished in: Neuroscience. 2008 Nov 19;157(2):484-94. doi: 10.1016/j.neuroscience.2008.07.050. PMID: 18555164 Free PMC article. Corrected and republished. Review.
Cited by
-
What Is the Role of Thalamostriatal Circuits in Learning Vocal Sequences?Front Neural Circuits. 2021 Sep 22;15:724858. doi: 10.3389/fncir.2021.724858. eCollection 2021. Front Neural Circuits. 2021. PMID: 34630047 Free PMC article.
-
Population coding of time-varying sounds in the non-lemniscal Inferior Colliculus.bioRxiv [Preprint]. 2023 Aug 16:2023.08.14.553263. doi: 10.1101/2023.08.14.553263. bioRxiv. 2023. Update in: J Neurophysiol. 2024 May 1;131(5):842-864. doi: 10.1152/jn.00013.2024. PMID: 37645904 Free PMC article. Updated. Preprint.
-
Reversible Inactivation of Ferret Auditory Cortex Impairs Spatial and Nonspatial Hearing.J Neurosci. 2023 Feb 1;43(5):749-763. doi: 10.1523/JNEUROSCI.1426-22.2022. Epub 2023 Jan 5. J Neurosci. 2023. PMID: 36604168 Free PMC article.
-
Effective Regulation of Auditory Processing by Parvalbumin Interneurons in the Tail of the Striatum.J Neurosci. 2024 Jan 31;44(5):e1171232023. doi: 10.1523/JNEUROSCI.1171-23.2023. J Neurosci. 2024. PMID: 38296650 Free PMC article.
-
[Neural mechanism for modulation of auditory response of the striatum by locomotion].Nan Fang Yi Ke Da Xue Xue Bao. 2022 May 20;42(5):766-771. doi: 10.12122/j.issn.1673-4254.2022.05.20. Nan Fang Yi Ke Da Xue Xue Bao. 2022. PMID: 35673923 Free PMC article. Chinese.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases