

Bimodal sensing of guidance cues in mechanically distinct microenvironments
- PMID: 30459308
- PMCID: PMC6244288
- DOI: 10.1038/s41467-018-07290-y
Bimodal sensing of guidance cues in mechanically distinct microenvironments
Abstract
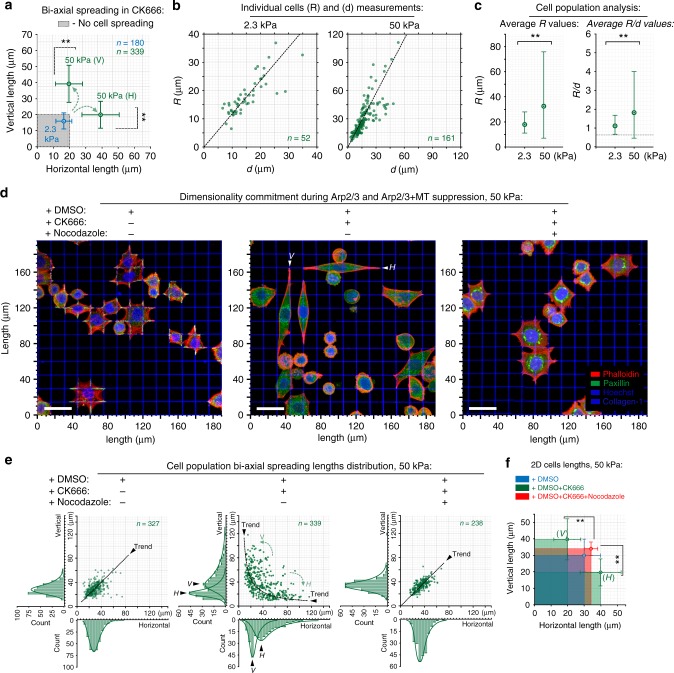
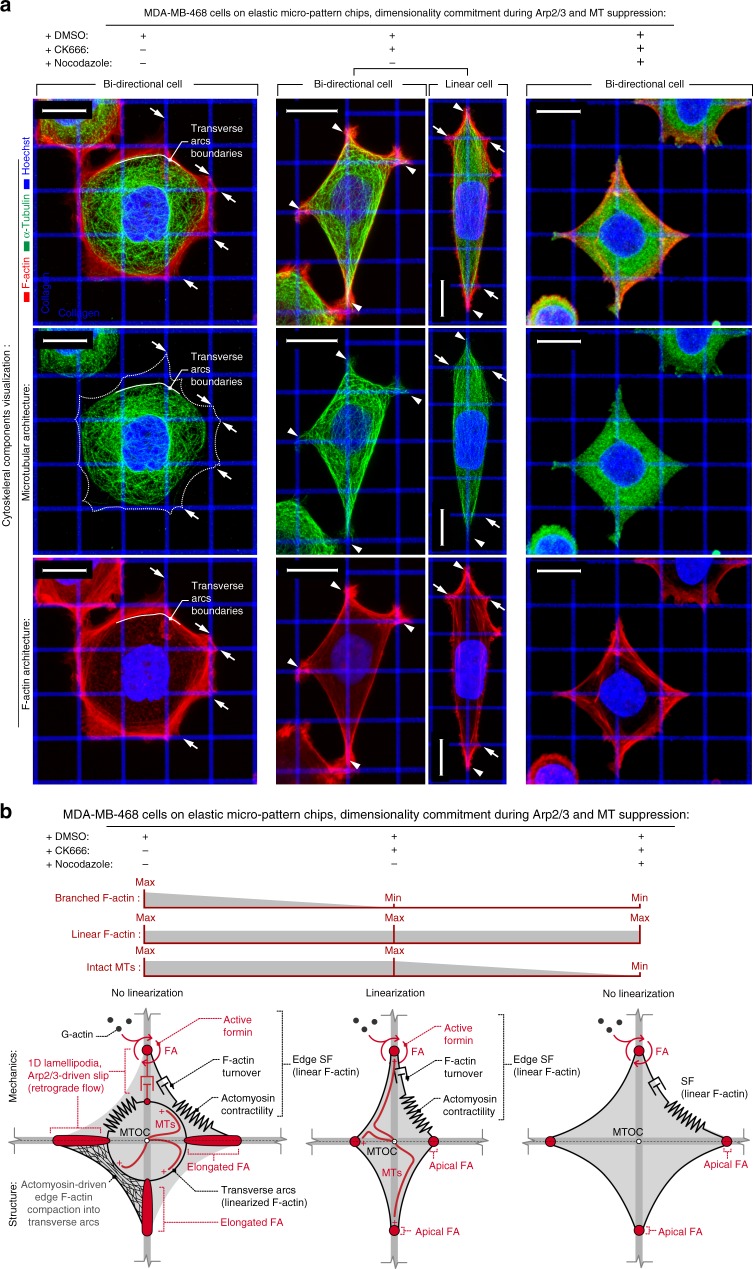
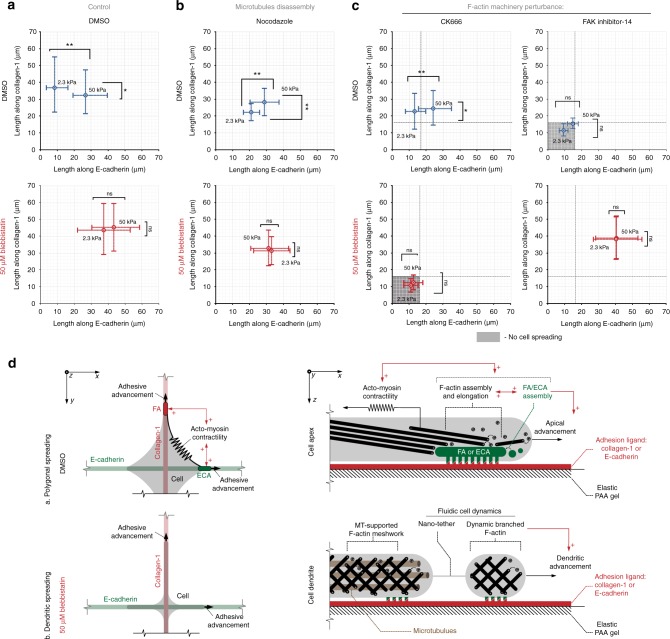
Contact guidance due to extracellular matrix architecture is a key regulator of carcinoma invasion and metastasis, yet our understanding of how cells sense guidance cues is limited. Here, using a platform with variable stiffness that facilitates uniaxial or biaxial matrix cues, or competing E-cadherin adhesions, we demonstrate distinct mechanoresponsive behavior. Through disruption of traction forces, we observe a profound phenotypic shift towards a mode of dendritic protrusion and identify bimodal processes that govern guidance sensing. In contractile cells, guidance sensing is strongly dependent on formins and FAK signaling and can be perturbed by disrupting microtubule dynamics, while low traction conditions initiate fluidic-like dendritic protrusions that are dependent on Arp2/3. Concomitant disruption of these bimodal mechanisms completely abrogates the contact guidance response. Thus, guidance sensing in carcinoma cells depends on both environment architecture and mechanical properties and targeting the bimodal responses may provide a rational strategy for disrupting metastatic behavior.
Conflict of interest statement
The authors declare no competing interests.
Figures
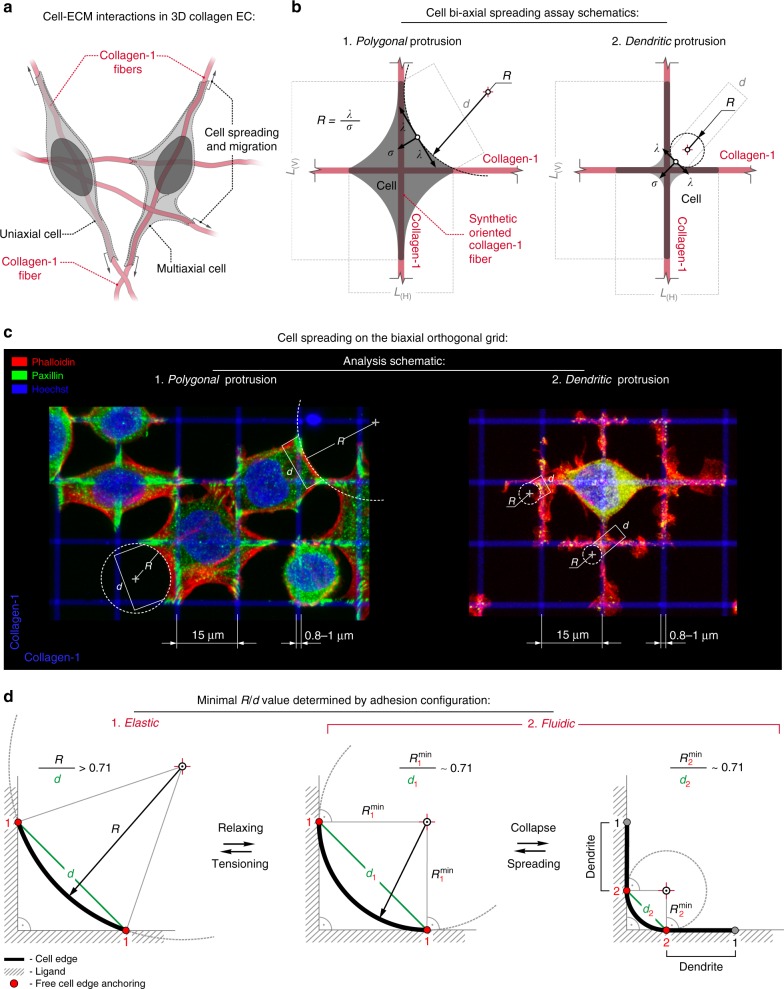
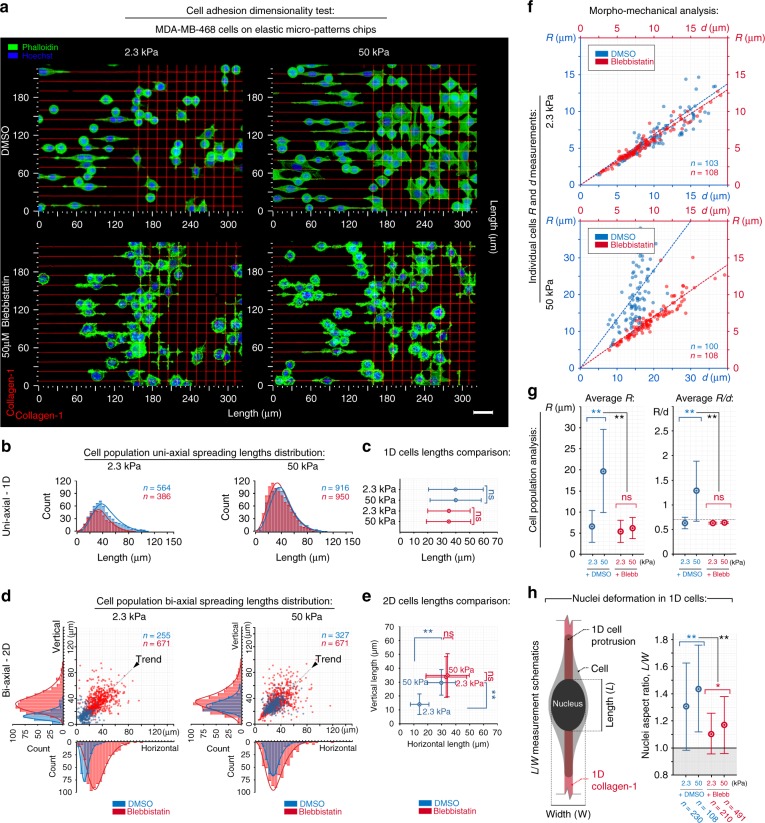
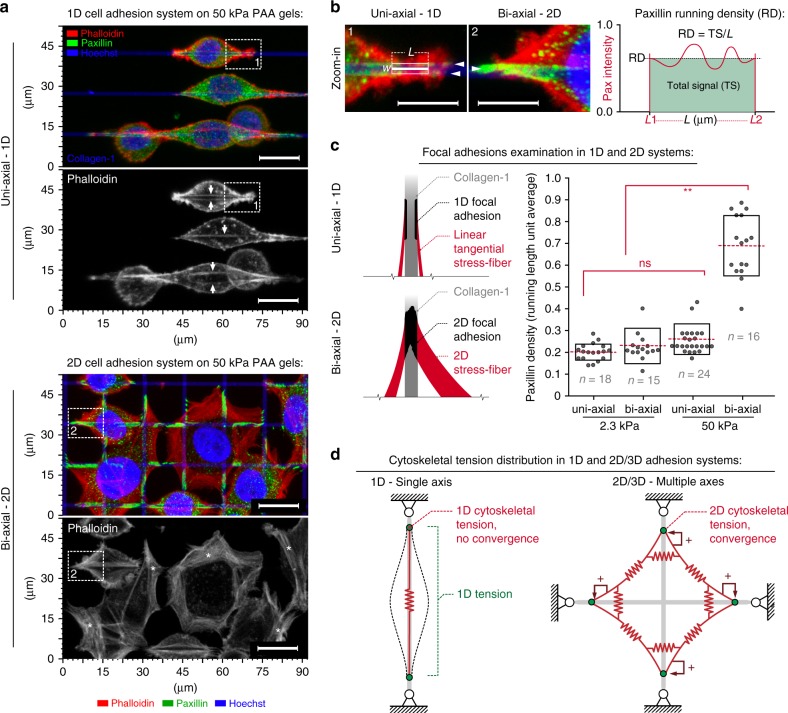
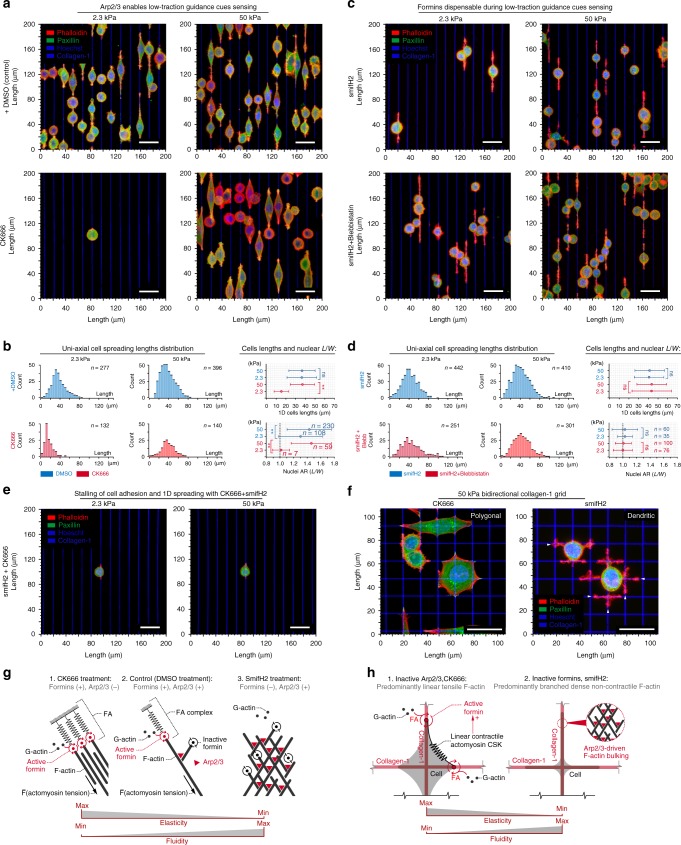

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
