

Rhythms of life: circadian disruption and brain disorders across the lifespan
- PMID: 30459365
- PMCID: PMC6338075
- DOI: 10.1038/s41583-018-0088-y
Rhythms of life: circadian disruption and brain disorders across the lifespan
Abstract
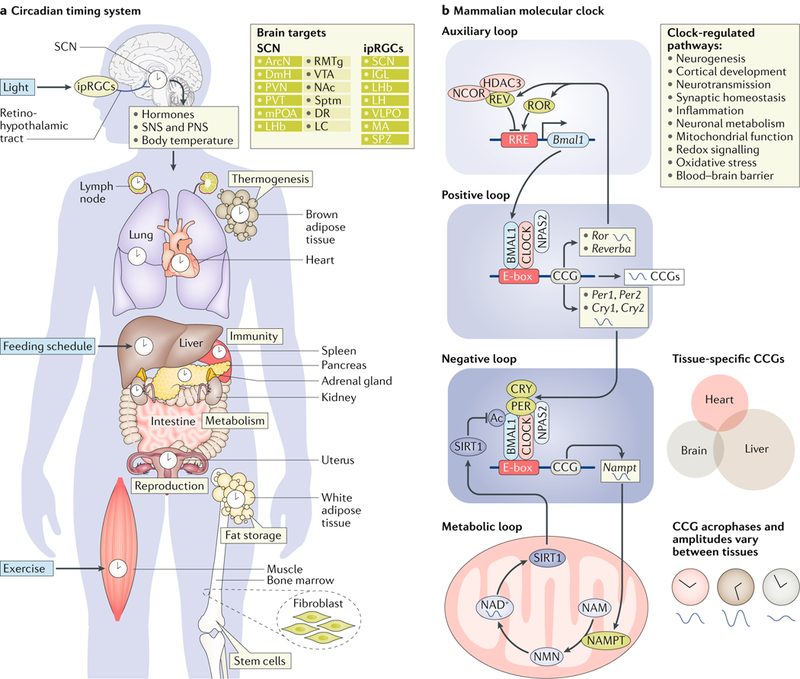
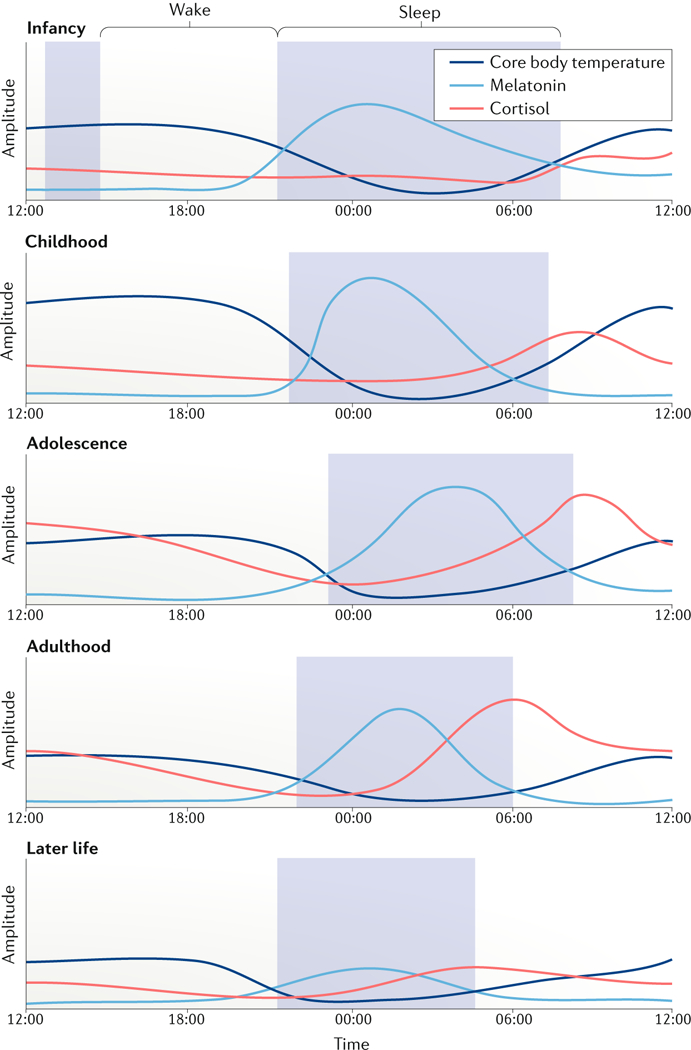
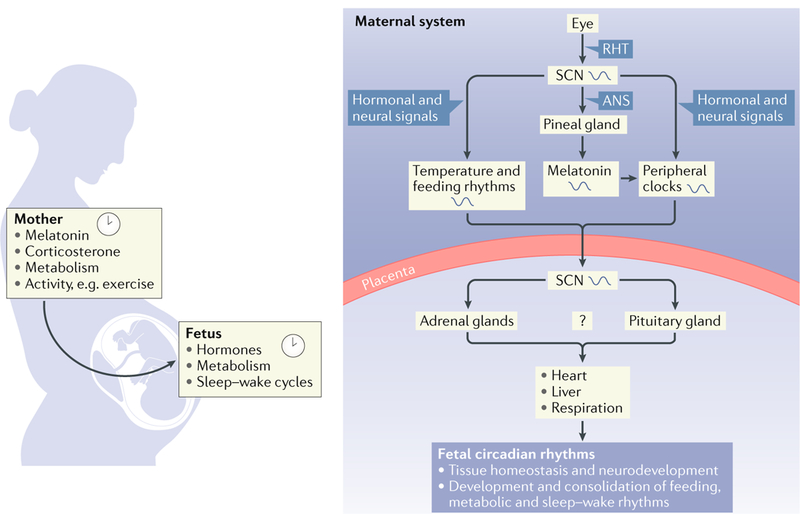
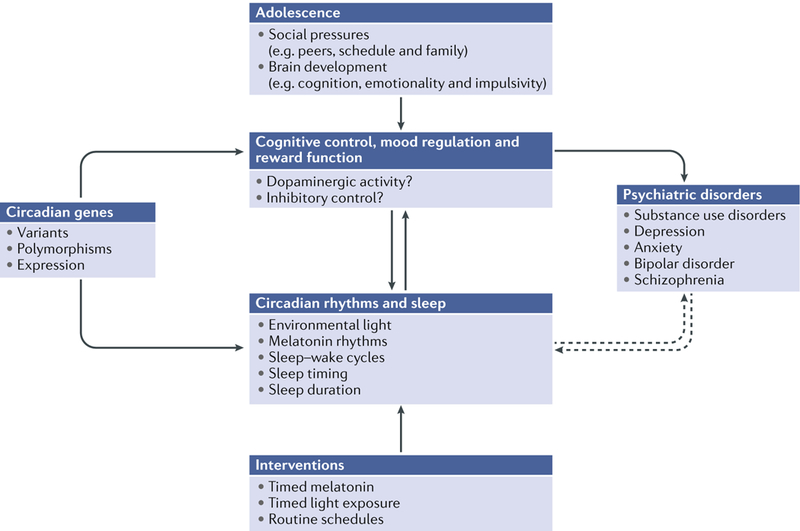
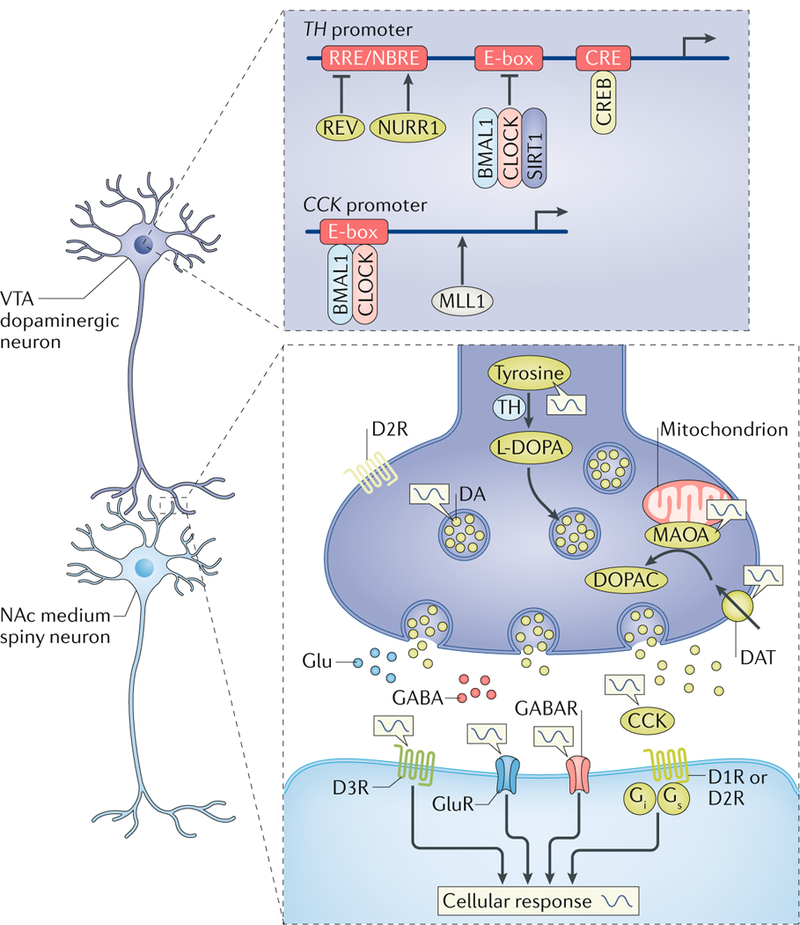
Many processes in the human body - including brain function - are regulated over the 24-hour cycle, and there are strong associations between disrupted circadian rhythms (for example, sleep-wake cycles) and disorders of the CNS. Brain disorders such as autism, depression and Parkinson disease typically develop at certain stages of life, and circadian rhythms are important during each stage of life for the regulation of processes that may influence the development of these disorders. Here, we describe circadian disruptions observed in various brain disorders throughout the human lifespan and highlight emerging evidence suggesting these disruptions affect the brain. Currently, much of the evidence linking brain disorders and circadian dysfunction is correlational, and so whether and what kind of causal relationships might exist are unclear. We therefore identify remaining questions that may direct future research towards a better understanding of the links between circadian disruption and CNS disorders.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures
References
-
- Reppert SM & Weaver DR Coordination of circadian timing in mammals. Nature 418, 935–941 (2002). - PubMed
-
- Mighdoll MI, Tao R, Kleinman JE & Hyde TM Myelin, myelin-related disorders, and psychosis. Schizophr. Res 161, 85–93 (2015). - PubMed
-
- Hsu PK, Ptacek LJ & Fu YH Genetics of human sleep behavioral phenotypes. Methods Enzymol 552, 309–324 (2015). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
