

Activation Stoichiometry and Pore Architecture of TRPA1 Probed with Channel Concatemers
- PMID: 30459425
- PMCID: PMC6244152
- DOI: 10.1038/s41598-018-35435-y
Activation Stoichiometry and Pore Architecture of TRPA1 Probed with Channel Concatemers
Abstract
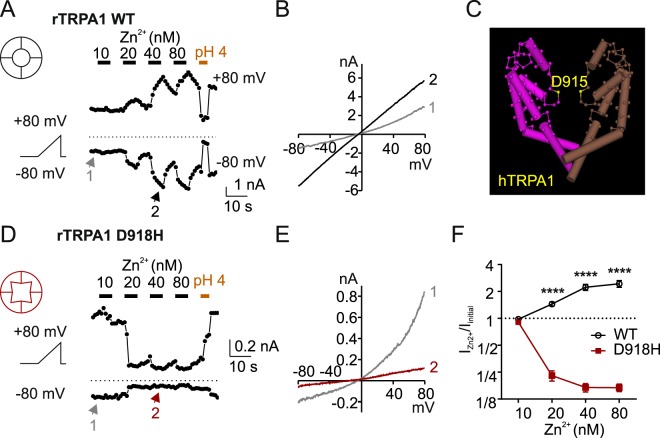
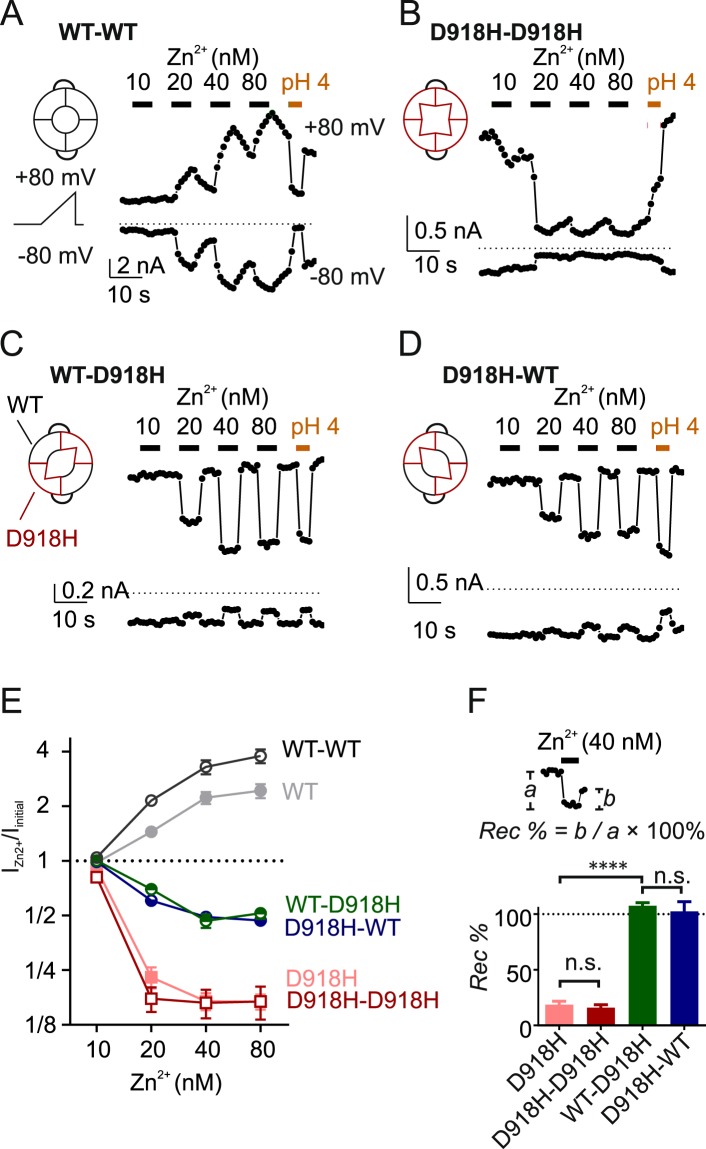
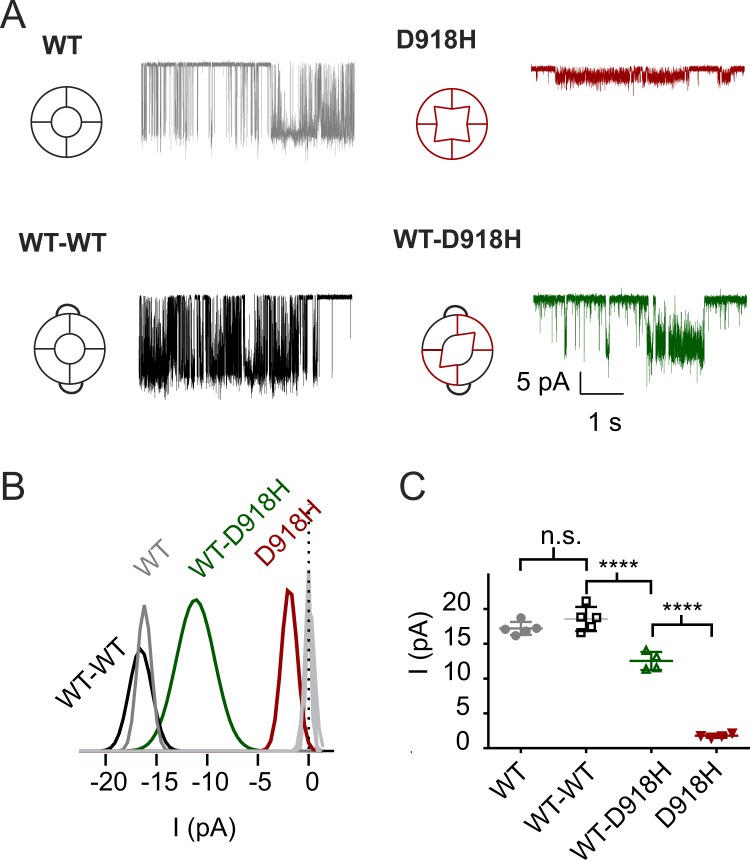
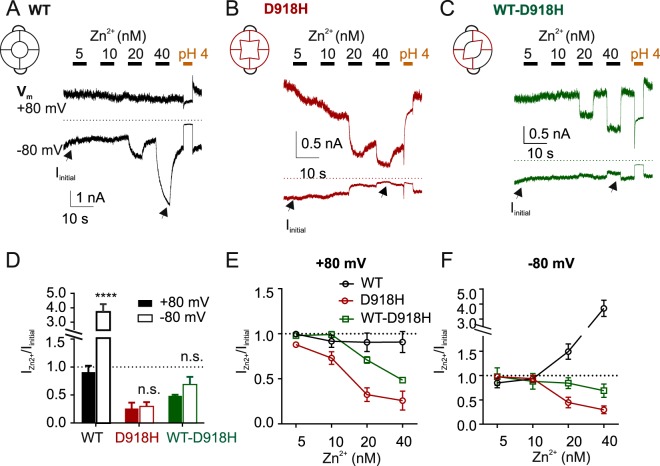
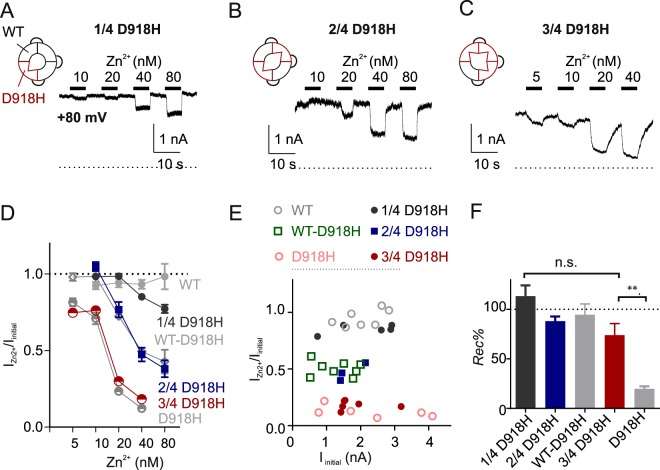
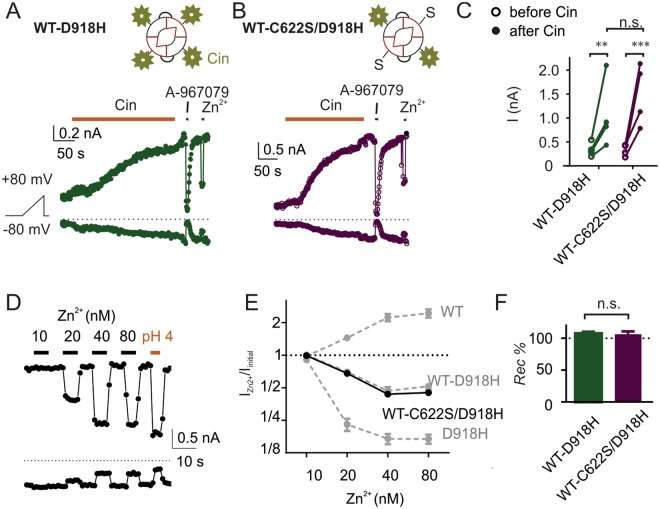
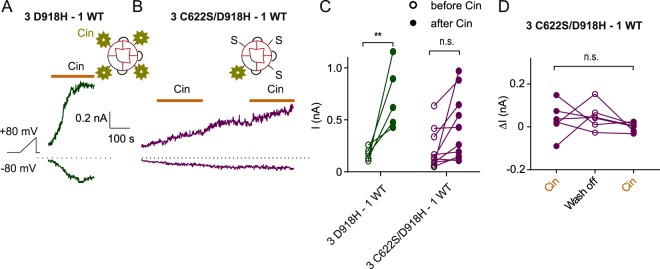
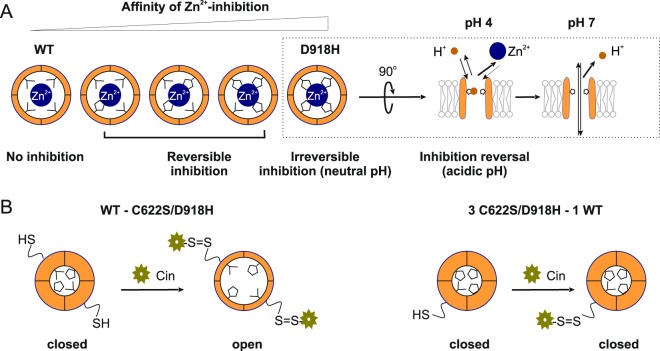
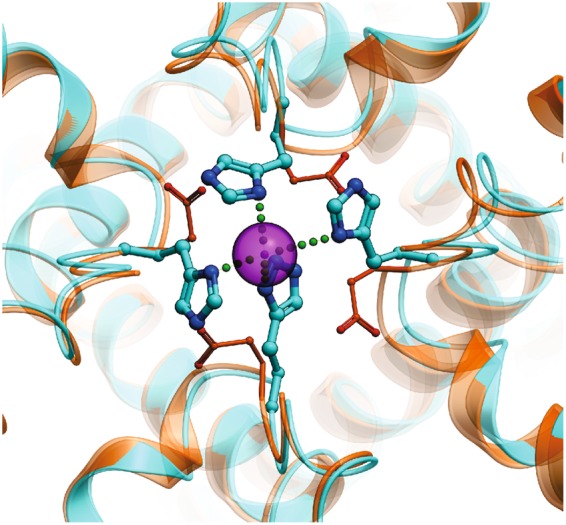
The nociceptor ion channel TRPA1 detects a wide range of hazardous chemicals, including reactive electrophiles such as cinnamaldehyde, which gate the channel allowing Na+ and Ca2+ entry. TRPA1 assembles as a tetramer, with a central pore within which an aspartate residue (D918) determines Ca2+ permeability. Here, we report that introduction of histidine at this position, D918H, makes TRPA1 channels sensitive to block by nanomolar concentration of Zn2+ and can be used to functionally tag subunits in concatemers. Concatemers with increasing numbers of D918H subunits display increasing sensitivity to Zn2+ inhibition, indicating that the four side chains at position 918 of the tetramer directly coordinate Zn2+ and other permeating divalent cations. In the published structure of TRPA1, this requires a rearrangement of the pore region which may represent the true open state of the channel. Concatemeric channels containing subunits mutated to be insensitive to reactive electrophiles (C622S) could be activated by cinnamaldehyde when as few as two subunits contained intact ligand binding sites. Activation upon liganding of just two of the four possible subunits may represent an optimal strategy to rapidly and reliably detect noxious chemicals.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- R01 DC013741/DC/NIDCD NIH HHS/United States
- R21 NS087633/NS/NINDS NIH HHS/United States
- R21NS087633/U.S. Department of Health & Human Services | NIH | National Institute of Neurological Disorders and Stroke (NINDS)/International
- R01DC013741/U.S. Department of Health & Human Services | NIH | National Institute on Deafness and Other Communication Disorders (NIDCD)/International
LinkOut - more resources
Full Text Sources
Miscellaneous
