

Comparative Genomics of Wolbachia- Cardinium Dual Endosymbiosis in a Plant-Parasitic Nematode
- PMID: 30459726
- PMCID: PMC6232779
- DOI: 10.3389/fmicb.2018.02482
Comparative Genomics of Wolbachia- Cardinium Dual Endosymbiosis in a Plant-Parasitic Nematode
Abstract
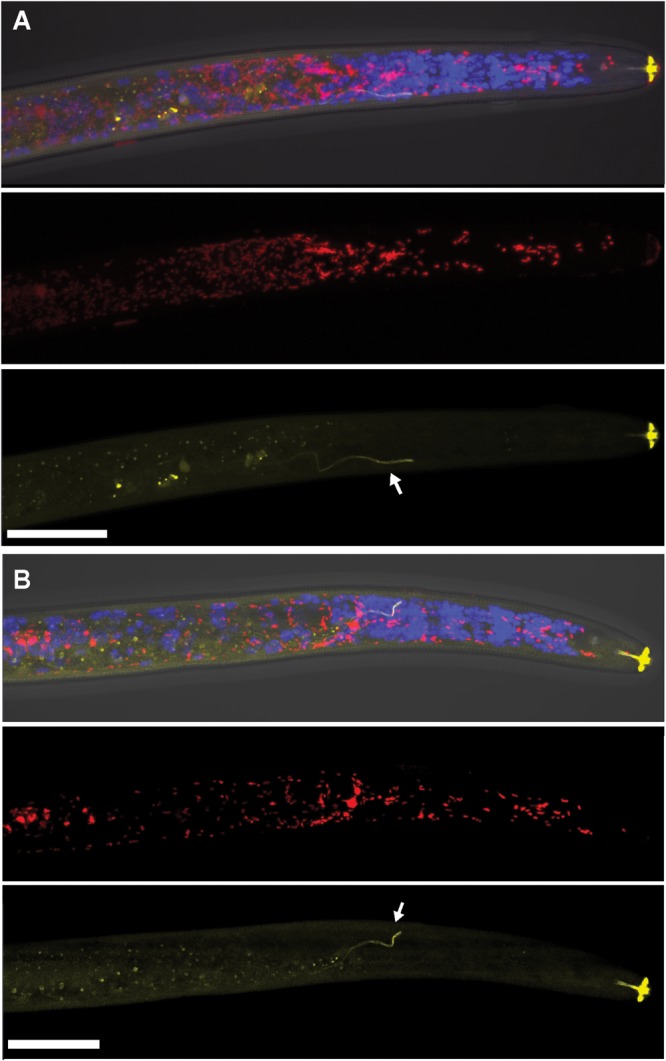
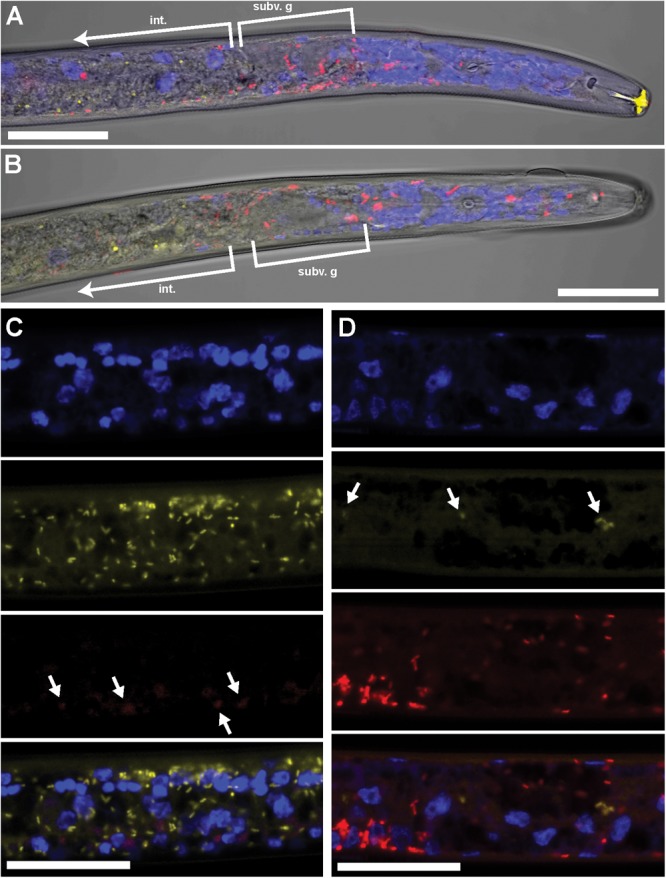
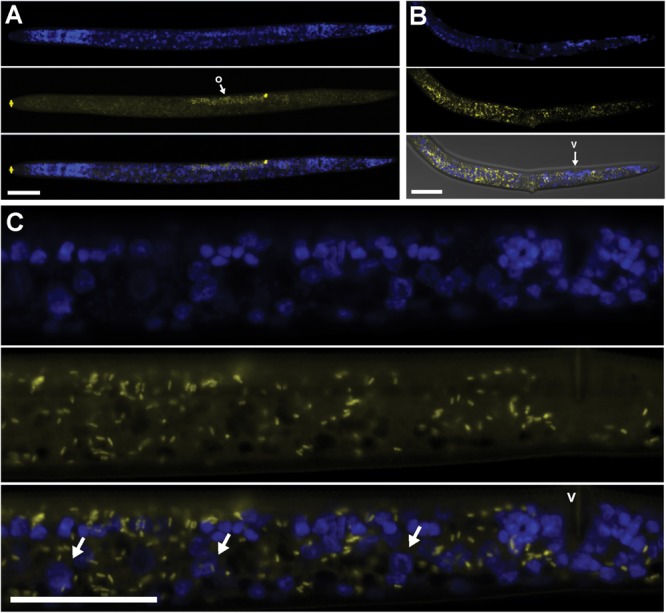
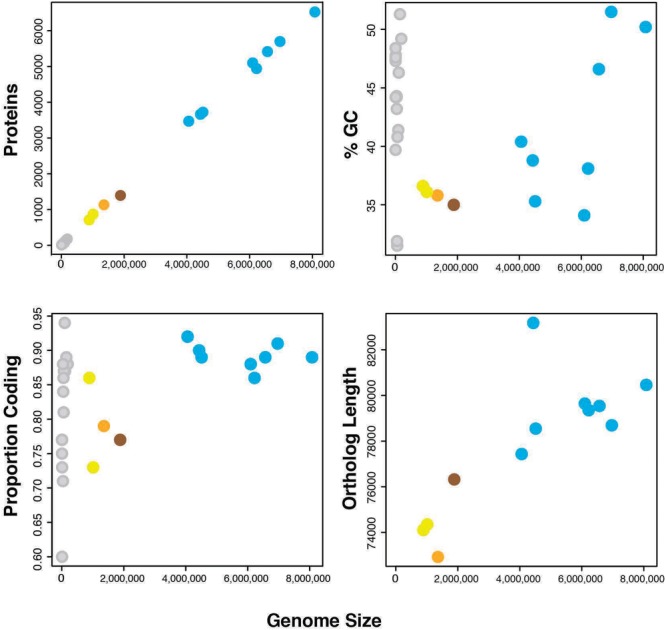
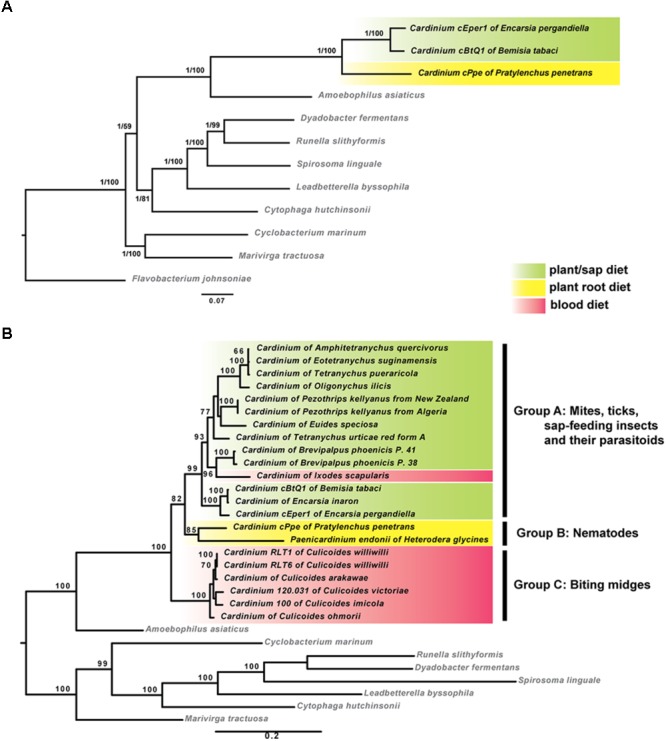
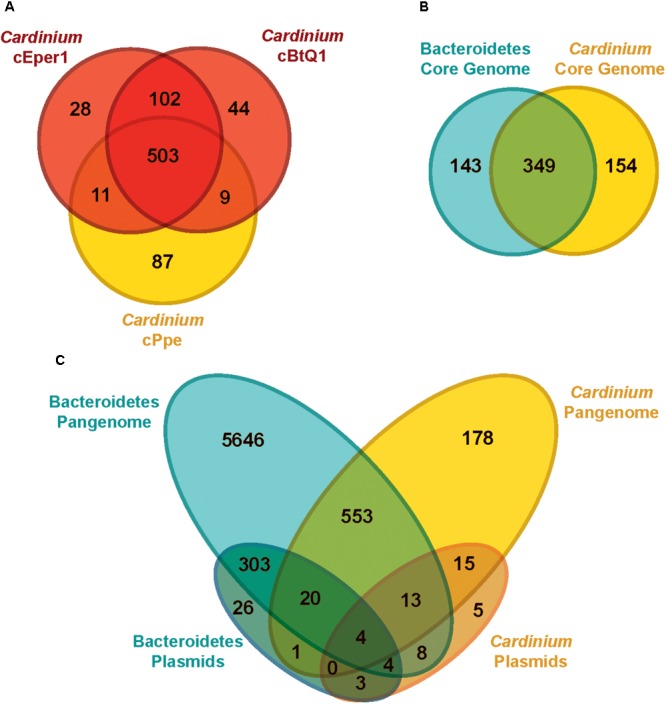
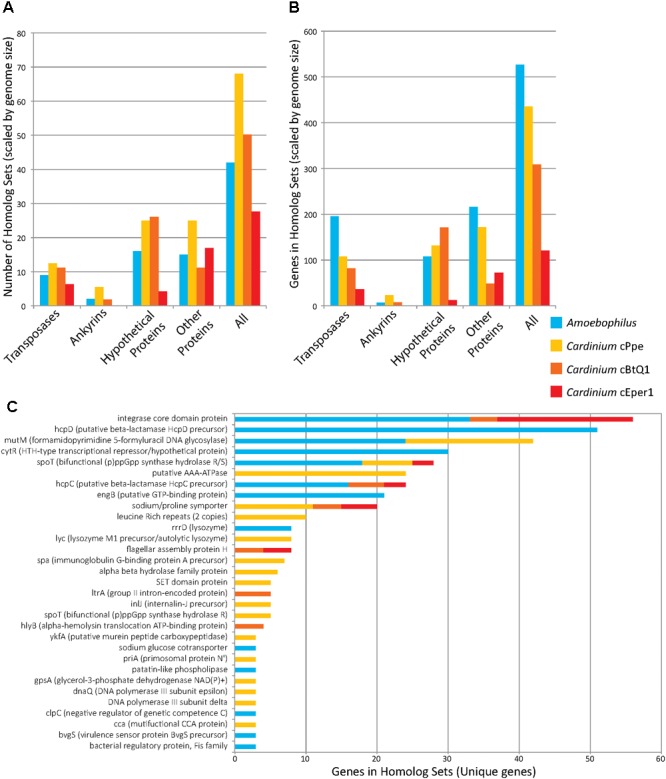
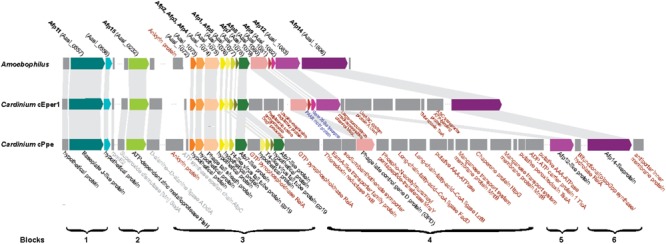
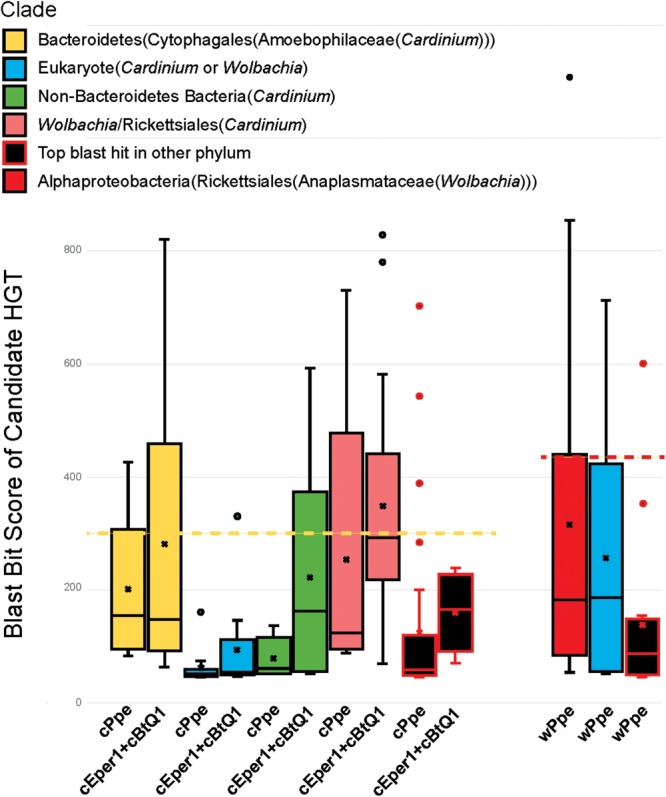
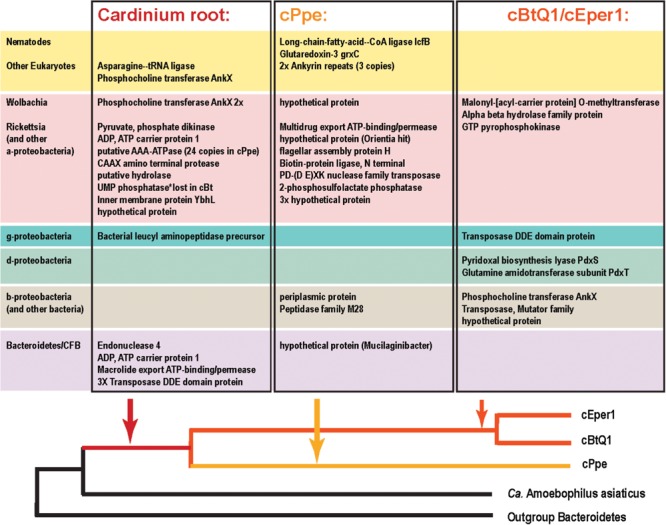
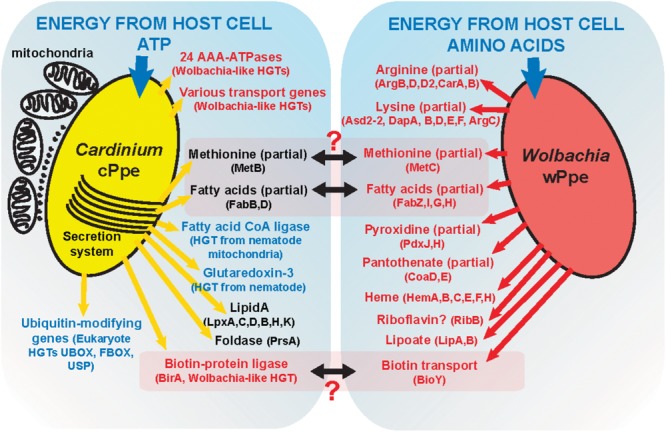
Wolbachia and Cardinium are among the most important and widespread of all endosymbionts, occurring in nematodes and more than half of insect and arachnid species, sometimes as coinfections. These symbionts are of significant interest as potential biocontrol agents due to their abilities to cause major effects on host biology and reproduction through cytoplasmic incompatibility, sex ratio distortion, or obligate mutualism. The ecological and metabolic effects of coinfections are not well understood. This study examined a Wolbachia-Cardinium coinfection in the plant-parasitic nematode (PPN), Pratylenchus penetrans, producing the first detailed study of such a coinfection using fluorescence in situ hybridization (FISH), polymerase chain reaction (PCR), and comparative genomic analysis. Results from FISH and single-nematode PCR showed 123/127 individuals in a focal population carried Cardinium (denoted strain cPpe), and 48% were coinfected with Wolbachia strain wPpe. Both endosymbionts showed dispersed tissue distribution with highest densities in the anterior intestinal walls and gonads. Phylogenomic analyses confirmed an early place of cPpe and long distance from a sister strain in another PPN, Heterodera glycines, supporting a long history of both Cardinium and Wolbachia in PPNs. The genome of cPpe was 1.36 Mbp with 35.8% GC content, 1,131 predicted genes, 41% having no known function, and missing biotin and lipoate synthetic capacity and a plasmid present in other strains, despite having a slightly larger genome compared to other sequenced Cardinium. The larger genome revealed expansions of gene families likely involved in host-cellular interactions. More than 2% of the genes of cPpe and wPpe were identified as candidate horizontally transferred genes, with some of these from eukaryotes, including nematodes. A model of the possible Wolbachia-Cardinium interaction is proposed with possible complementation in function for pathways such as methionine and fatty acid biosynthesis and biotin transport.
Keywords: Cardinium; Pratylenchus penetrans; Wolbachia; endosymbiont; genomics; horizontal gene transfer; plant-parasitic nematode; symbiosis.
Figures
References
-
- An P. N. T., Yamaguchi M., Fukusaki E. (2017). Metabolic profiling of Drosophila melanogaster metamorphosis: a new insight into the central metabolic pathways. Metabolomics 13 1–13. 10.1007/s11306-017-1167-1 - DOI
LinkOut - more resources
Full Text Sources
Miscellaneous