

Disruption of cardiac Med1 inhibits RNA polymerase II promoter occupancy and promotes chromatin remodeling
- PMID: 30461303
- PMCID: PMC6397381
- DOI: 10.1152/ajpheart.00580.2018
Disruption of cardiac Med1 inhibits RNA polymerase II promoter occupancy and promotes chromatin remodeling
Abstract
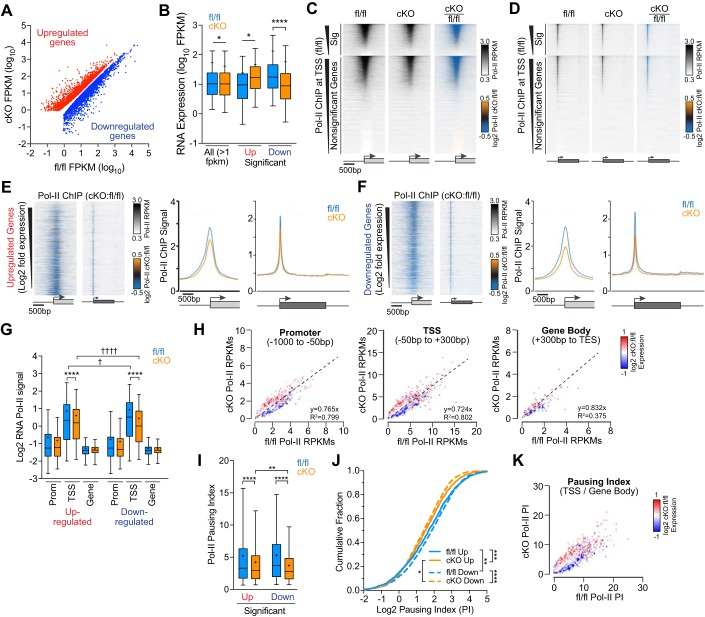
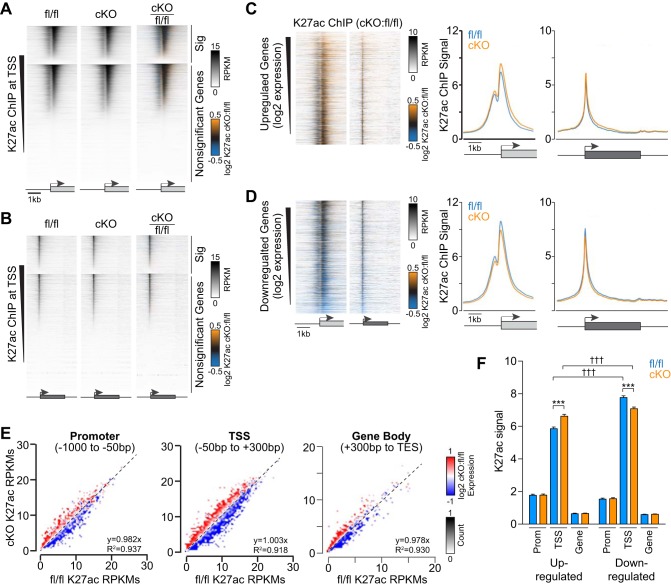
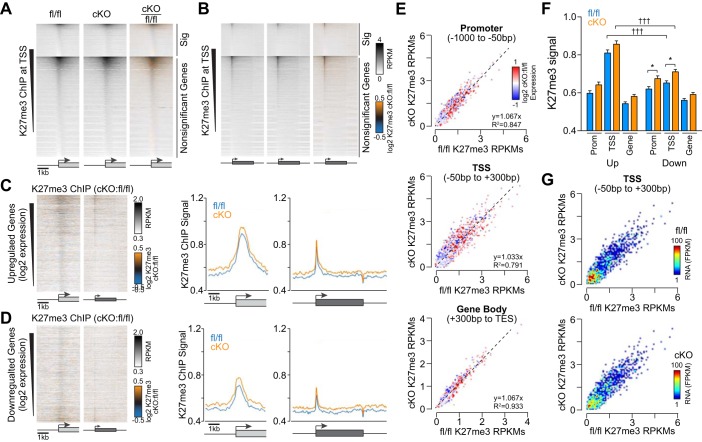
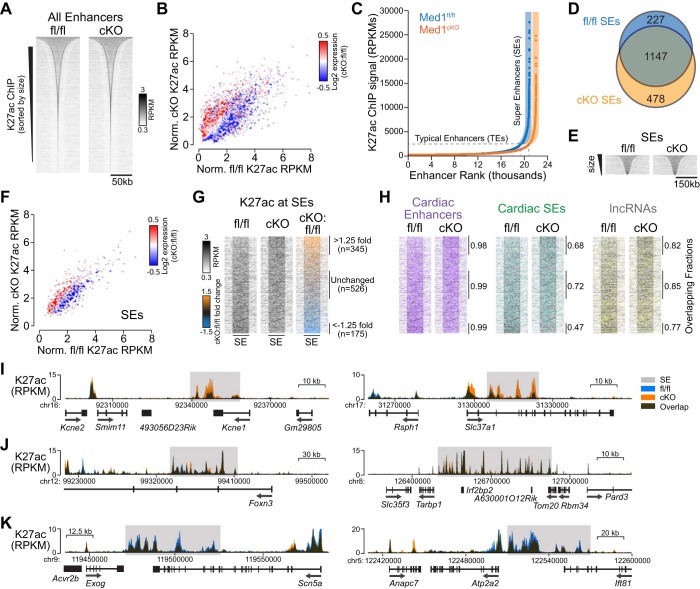
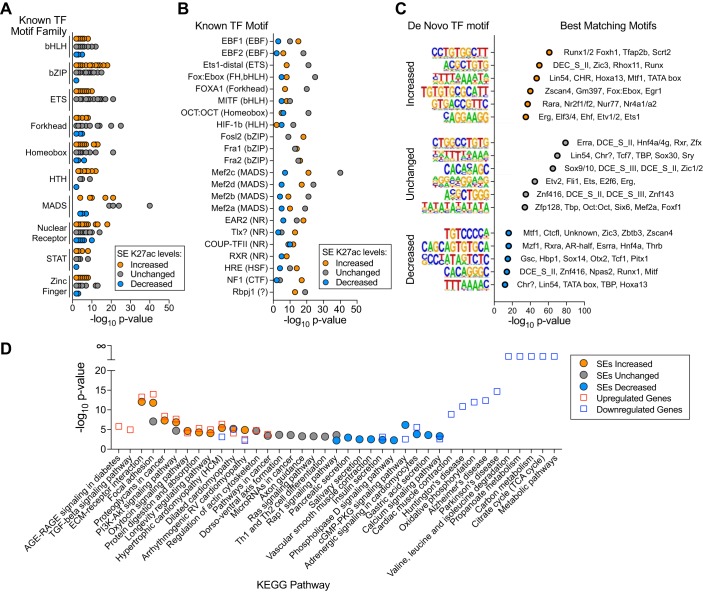
The Mediator coactivator complex directs gene-specific expression by binding distal enhancer-bound transcription factors through its Med1 subunit while bridging to RNA polymerase II (Pol II) at gene promoters. In addition, Mediator scaffolds epigenetic modifying enzymes that determine local DNA accessibility. Previously, we found that deletion of Med1 in cardiomyocytes deregulates more than 5,000 genes and promotes acute heart failure. Therefore, we hypothesized that Med1 deficiency disrupts enhancer-promoter coupling. Using chromatin immunoprecipitation-coupled deep sequencing (ChIP-seq; n = 3/ChIP assay), we found that the Pol II pausing index is increased in Med1 knockout versus floxed control mouse hearts primarily due to a decrease in Pol II occupancy at the majority of transcriptional start sites without a corresponding increase in elongating species. Parallel ChIP-seq assays reveal that Med1-dependent gene expression correlates strongly with histone H3 K27 acetylation, which is indicative of open and active chromatin at transcriptional start sites, whereas H3 K27 trimethylated levels, representing condensed and repressed DNA, are broadly increased and inversely correlate with absolute expression levels. Furthermore, Med1 deletion leads to dynamic changes in acetyl-K27 associated superenhancer regions and their enriched transcription factor-binding motifs that are consistent with altered gene expression. Our findings suggest that Med1 is important in establishing enhancer-promoter coupling in the heart and supports the proposed role of Mediator in establishing preinitiation complex formation. We also found that Med1 determines chromatin accessibility within genes and enhancer regions and propose that the composition of transcription factors associated with superenhancer changes to direct gene-specific expression. NEW & NOTEWORTHY Based on our previous findings that transcriptional homeostasis and cardiac function are disturbed by cardiomyocyte deletion of the Mediator coactivator Med1 subunit, we investigated potential underlying changes in RNA polymerase II localization and global chromatin accessibility. Using chromatin immunoprecipitation sequencing, we found that disrupted transcription arises from a deficit in RNA polymerase II recruitment to gene promoters. Furthermore, active versus repressive chromatin marks are redistributed within gene loci and at enhancer regions correlated with gene expression changes.
Keywords: RNA polymerase II; chromatin immunoprecipitation-coupled deep sequencing; enhancer; heart; histone H3 lysine 27; mediator.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the authors.
Figures
References
-
- Acevedo ML, Kraus WL. Mediator and p300/CBP-steroid receptor coactivator complexes have distinct roles, but function synergistically, during estrogen receptor alpha-dependent transcription with chromatin templates. Mol Cell Biol 23: 335–348, 2003. doi:10.1128/MCB.23.1.335-348.2003. - DOI - PMC - PubMed
-
- Anand P, Brown JD, Lin CY, Qi J, Zhang R, Artero PC, Alaiti MA, Bullard J, Alazem K, Margulies KB, Cappola TP, Lemieux M, Plutzky J, Bradner JE, Haldar SM. BET bromodomains mediate transcriptional pause release in heart failure. Cell 154: 569–582, 2013. doi:10.1016/j.cell.2013.07.013. - DOI - PMC - PubMed
-
- Aranda-Orgilles B, Saldaña-Meyer R, Wang E, Trompouki E, Fassl A, Lau S, Mullenders J, Rocha PP, Raviram R, Guillamot M, Sánchez-Díaz M, Wang K, Kayembe C, Zhang N, Amoasii L, Choudhuri A, Skok JA, Schober M, Reinberg D, Sicinski P, Schrewe H, Tsirigos A, Zon LI, Aifantis I. MED12 regulates HSC-specific enhancers independently of mediator kinase activity to control hematopoiesis. Cell Stem Cell 19: 784–799, 2016. doi:10.1016/j.stem.2016.08.004. - DOI - PMC - PubMed
-
- Baskin KK, Grueter CE, Kusminski CM, Holland WL, Bookout AL, Satapati S, Kong YM, Burgess SC, Malloy CR, Scherer PE, Newgard CB, Bassel-Duby R, Olson EN. MED13-dependent signaling from the heart confers leanness by enhancing metabolism in adipose tissue and liver. EMBO Mol Med 6: 1610–1621, 2014. doi:10.15252/emmm.201404218. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases