

Aminocyclopropane-1-carboxylic acid is a key regulator of guard mother cell terminal division in Arabidopsis thaliana
- PMID: 30462272
- PMCID: PMC6363092
- DOI: 10.1093/jxb/ery413
Aminocyclopropane-1-carboxylic acid is a key regulator of guard mother cell terminal division in Arabidopsis thaliana
Abstract
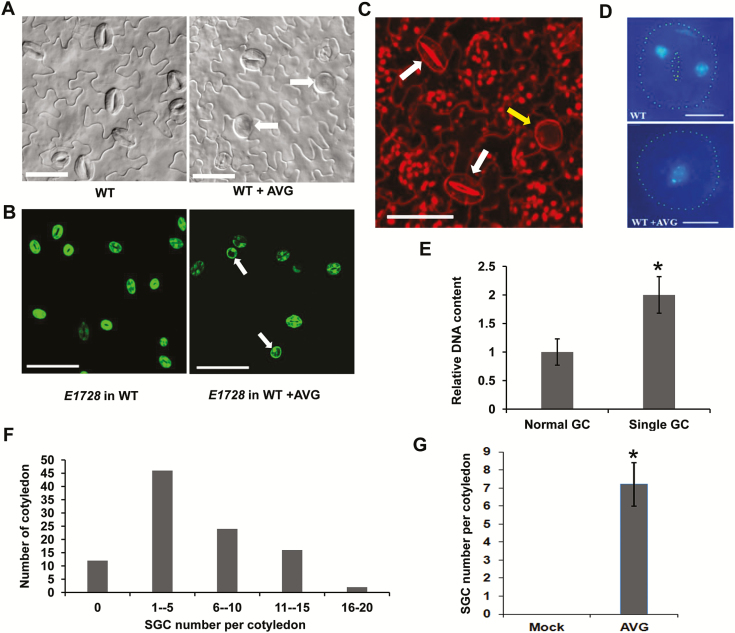
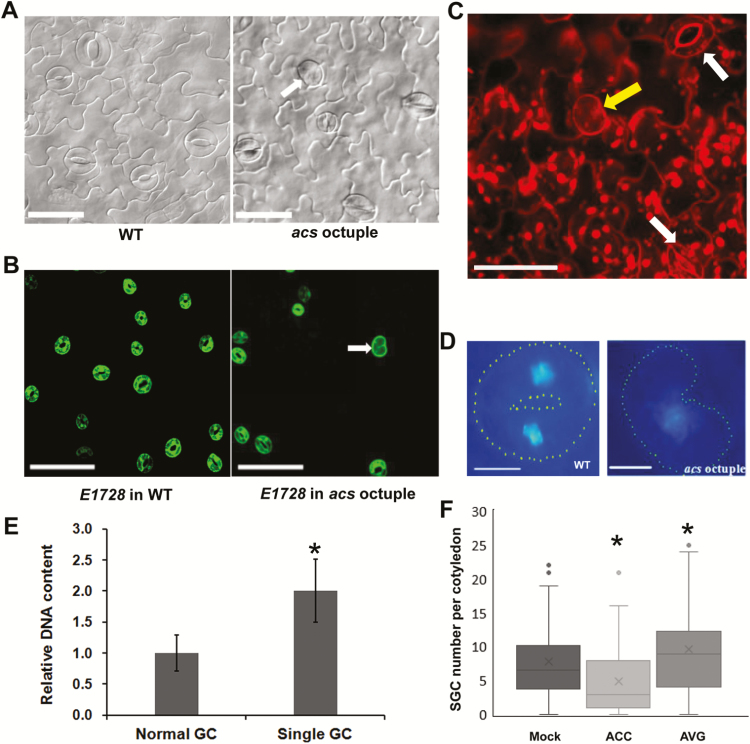
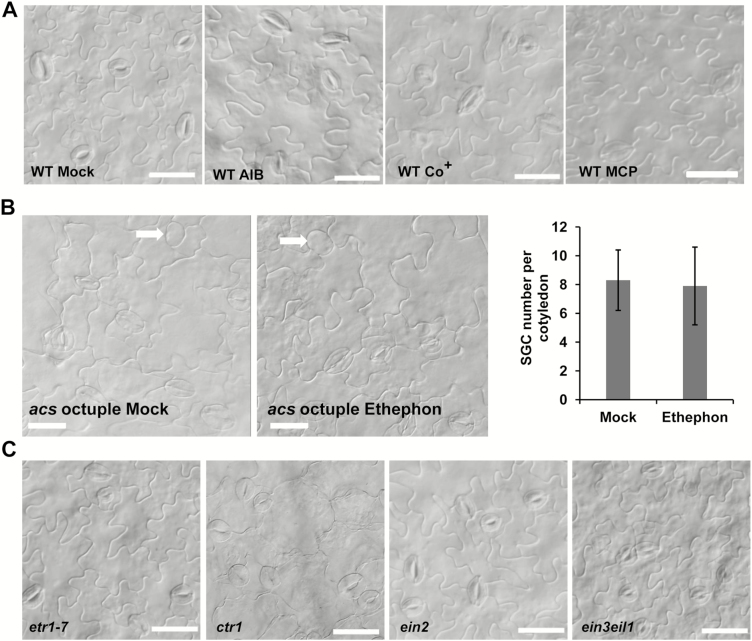
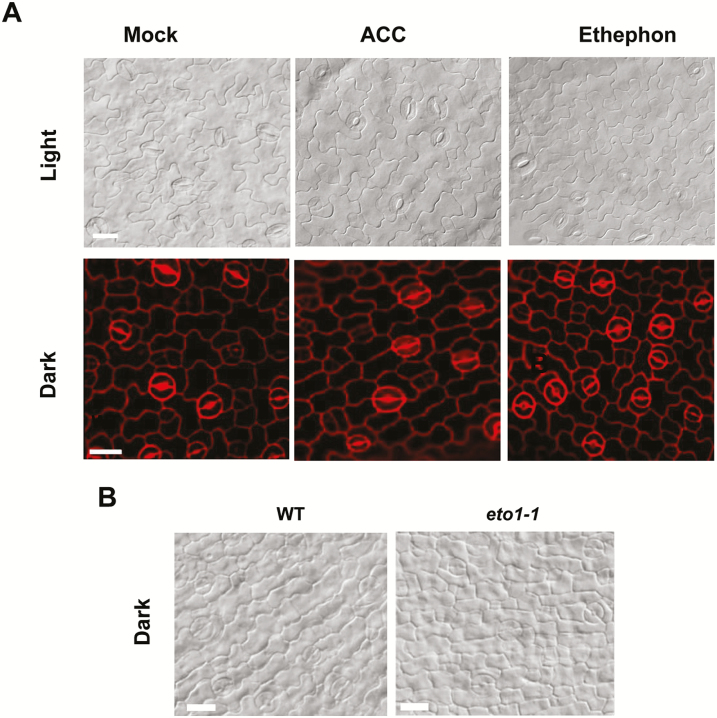
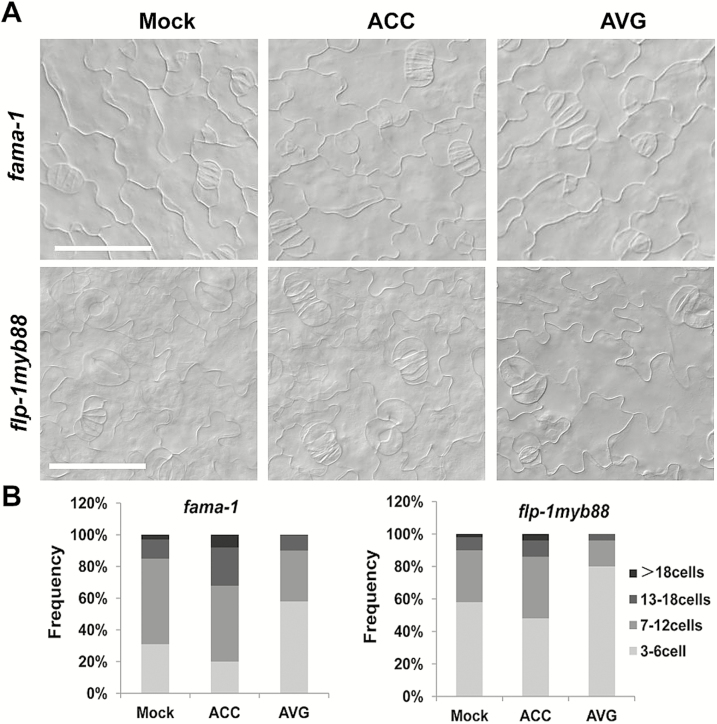
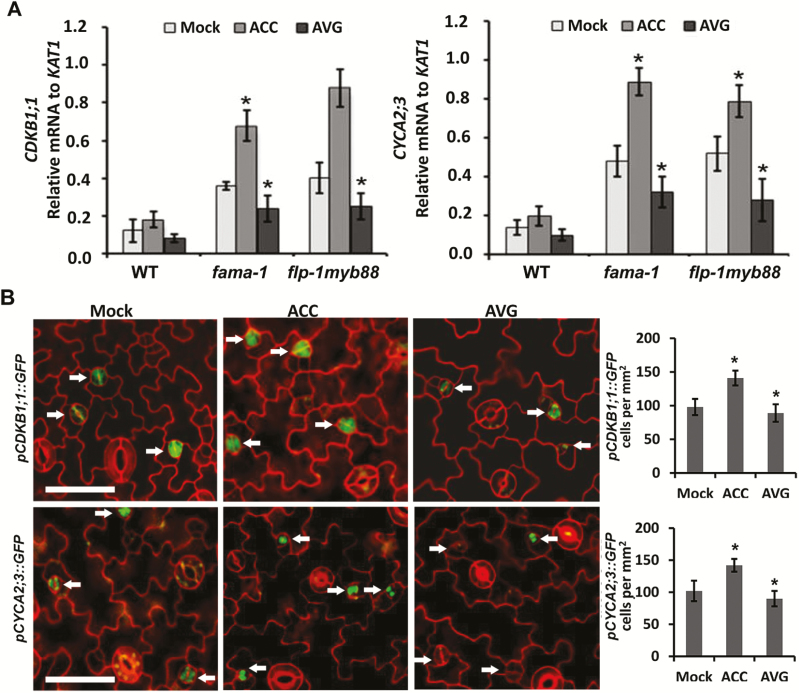
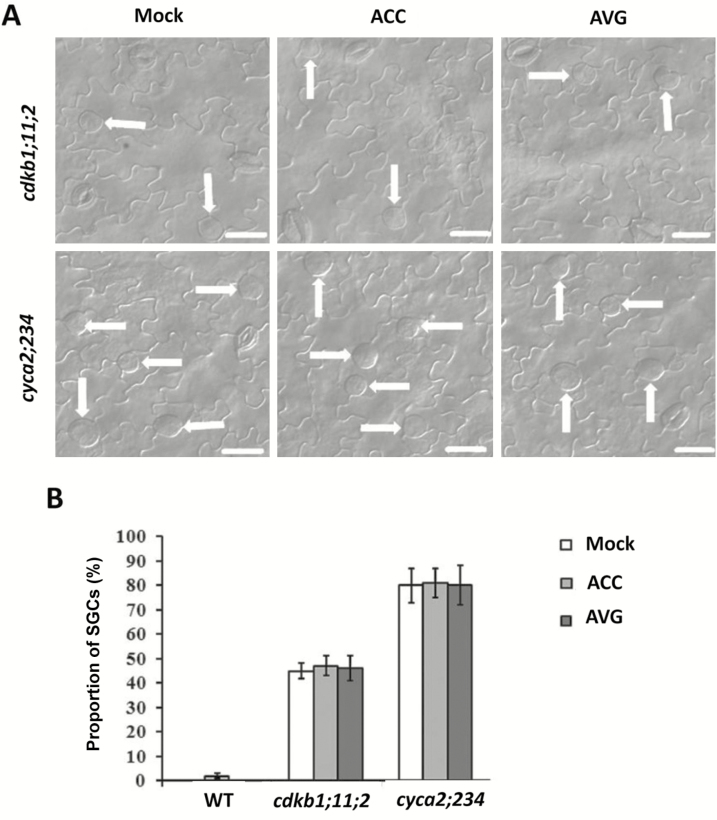
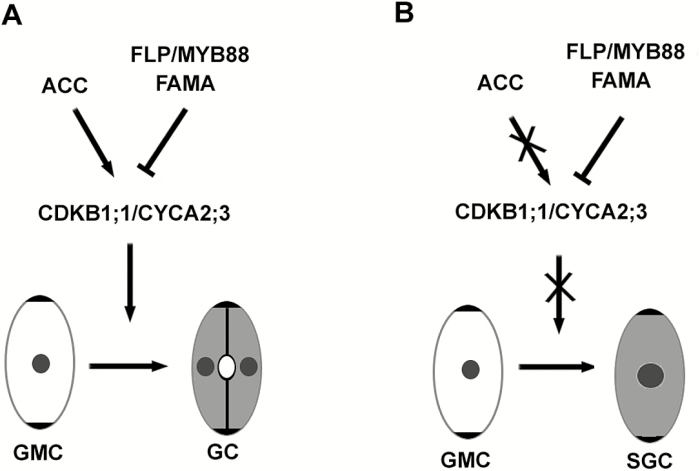
Stomata have a critical function in the exchange of gases and water vapor between plants and their environment. Stomatal development is under the rigorous control of many regulators. The last step of development is the terminal division of guard mother cells (GMC) into two guard cells (GC). It is still unclear how the symmetric division of GMCs is regulated. Here, we show that the ethylene precursor aminocyclopropane-1-carboxylic acid (ACC) is required for the symmetric division of GMCs into GCs in Arabidopsis. Exogenous application of the ACC biosynthesis inhibitor aminoethoxyvinylglycine (AVG) induced the formation of single guard cells (SGCs). Correspondingly, an acs octuple-mutant with extremely low endogenous ACC also developed SGCs, and exogenous ACC dramatically decreased the number of SGCs in this mutant whereas exogenous ethephon (which is gradually converted into ethylene) had no effect. Furthermore, neither blocking of endogenous ethylene synthesis nor disruption of ethylene signaling transduction could induce the production of SGCs. Further investigation indicated that ACC promoted the division of GMCs in fama-1 and flp-1myb88 mutants whereas AVG inhibited it. Moreover, ACC positively regulated the expression of CDKB1;1 and CYCA2;3 in the fama-1 and flp-1myb88 mutants. The SGC number was not affected by ACC or AVG in cdkb1;11;2 and cyca2;234 mutants. Taken together, the results demonstrate that ACC itself, but not ethylene, positively modulates the symmetric division of GMCs in a manner that is dependent on CDKB1s and CYCA2s.
Figures
References
-
- Alonso JM, Hirayama T, Roman G, Nourizadeh S, Ecker JR. 1999. EIN2, a bifunctional transducer of ethylene and stress responses in Arabidopsis. Science 284, 2148–2152. - PubMed
-
- Balcerowicz M, Ranjan A, Rupprecht L, Fiene G, Hoecker U. 2014. Auxin represses stomatal development in dark-grown seedlings via Aux/IAA proteins. Development 141, 3165–3176. - PubMed
-
- Bergmann DC, Sack FD. 2007. Stomatal development. Annual Review of Plant Biology 58, 163–181. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
