

Finely tuned conformational dynamics regulate the protective function of the lncRNA MALAT1 triple helix
- PMID: 30462290
- PMCID: PMC6379651
- DOI: 10.1093/nar/gky1171
Finely tuned conformational dynamics regulate the protective function of the lncRNA MALAT1 triple helix
Abstract
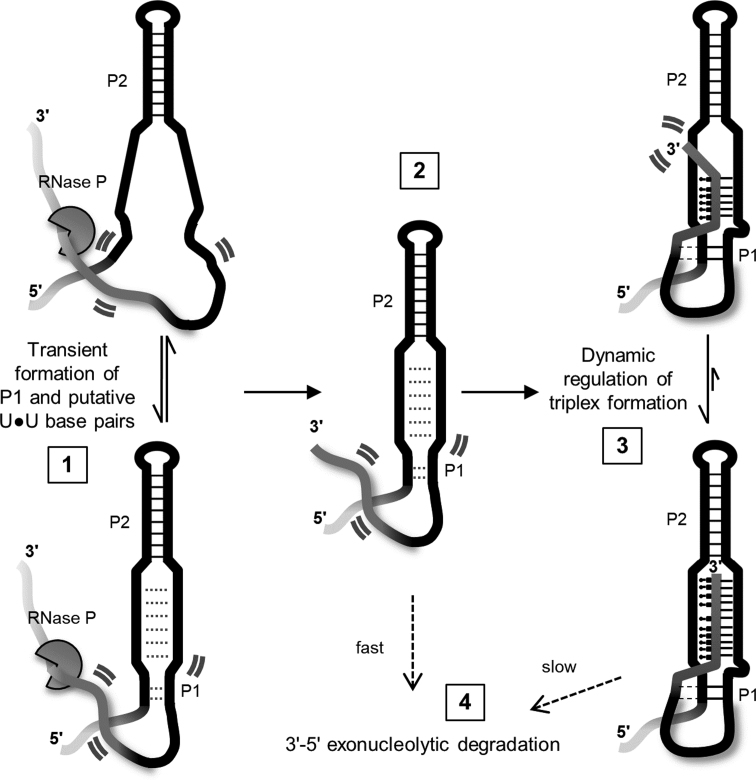
Nucleic acid triplexes may regulate many important biological processes. Persistent accumulation of the oncogenic 7-kb long noncoding RNA MALAT1 is dependent on an unusually long intramolecular triple helix. This triplex structure is positioned within a conserved ENE (element for nuclear expression) motif at the lncRNA 3' terminus and protects the entire transcript from degradation in a polyA-independent manner. A requisite 3' maturation step leads to triplex formation though the precise mechanism of triplex folding remains unclear. Furthermore, the contributions of several peripheral structural elements to triplex formation and protective function have not been determined. We evaluated the stability, conformational fluctuations, and function of this MALAT1 ENE triple helix (M1TH) protective element using in vitro mutational analyses coupled with biochemical and biophysical characterizations. Using fluorescence and UV melts, FRET, and an exonucleolytic decay assay we define a concerted mechanism for triplex formation and uncover a metastable, dynamic triplex population under near-physiological conditions. Structural elements surrounding the triplex regulate the dynamic M1TH conformational variability, but increased triplex dynamics lead to M1TH degradation. Taken together, we suggest that finely tuned dynamics may be a general mechanism regulating triplex-mediated functions.
© The Author(s) 2018. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
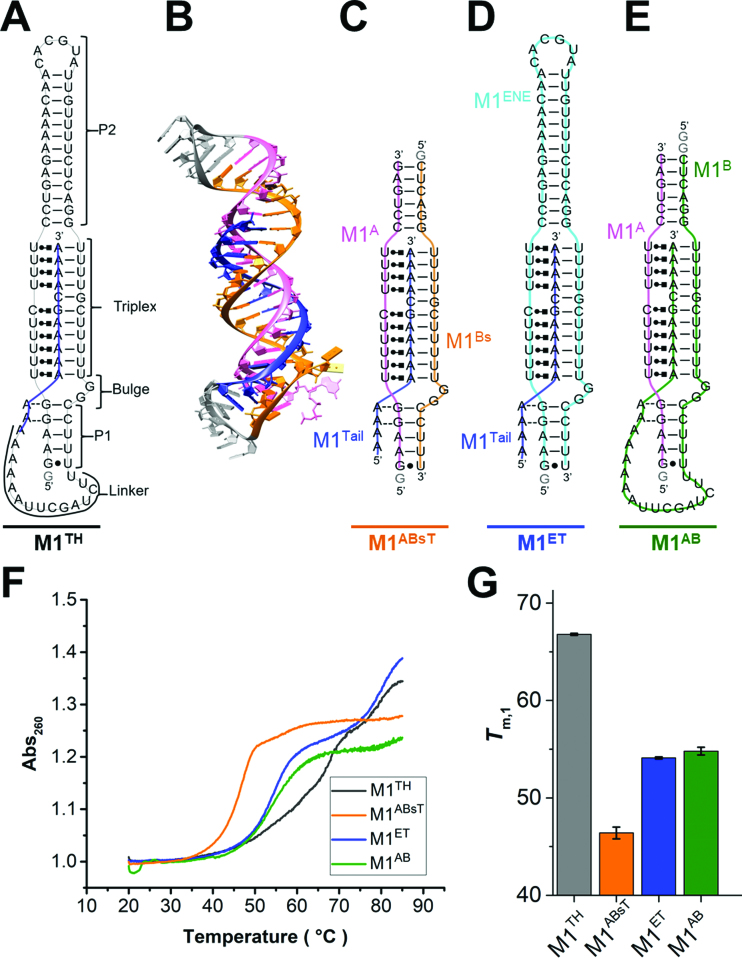
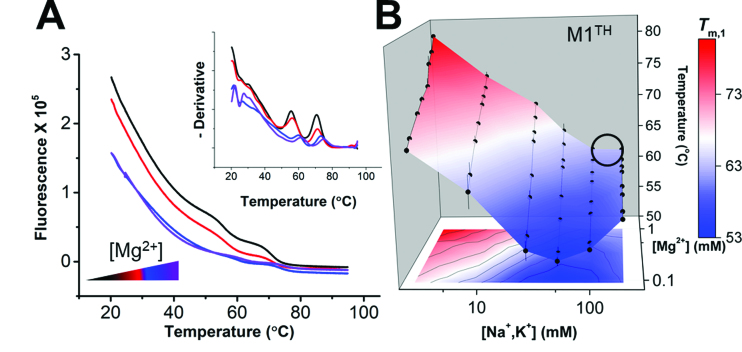
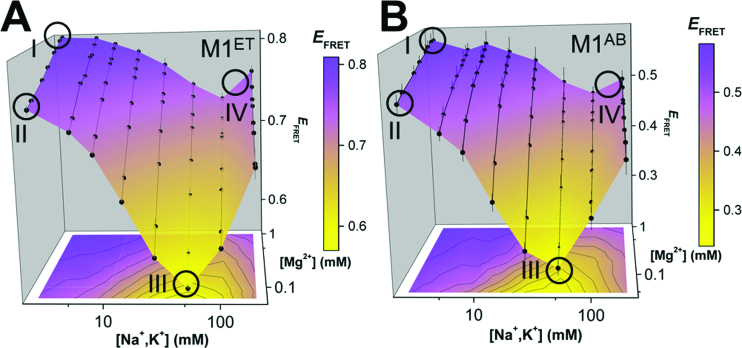
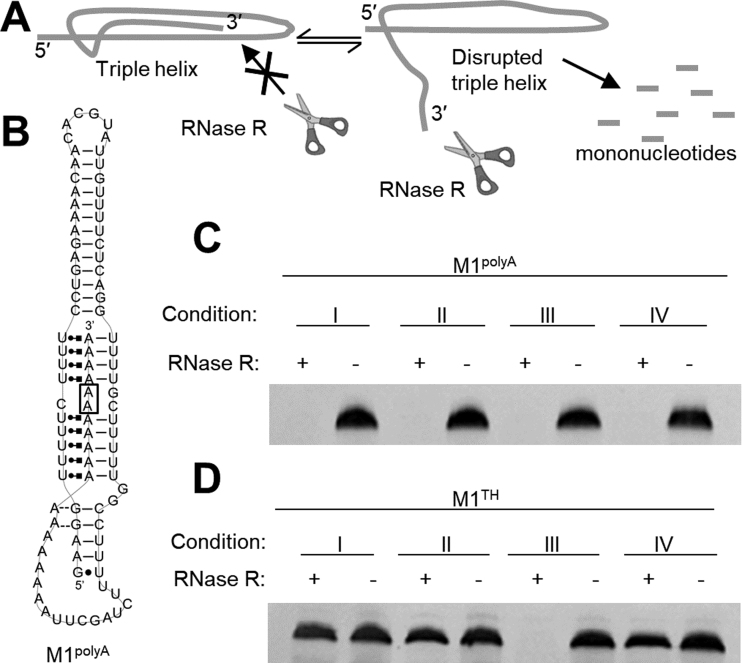
 ), A-minor interactions (−−), and a wobble base pair (•). A 5′ GTP (grey) is added to the sequence to facilitate transcription. The A-rich tail is colored blue and structural regions are indicated in brackets. (B) The crystal structure of the MALAT1 triple helix core (PDB: 4PLX) (33). The three strands are colored orange, pink, and blue. The two short loops (gray) were engineered to facilitate crystallization. (C) Secondary structure of trimolecular M1ABsT. M1Bs is colored orange, M1A is colored pink, and M1Tail is colored blue, as in B. (D) Secondary structure of bimolecular M1ET, which eliminates the basal linker. The ENE motif is colored cyan and the M1Tail is colored as in C. (E) Secondary structure of bimolecular M1AB, wherein the basal linker is included and the apical P2 helix is truncated. M1A is colored as in C and M1B is colored green. In C–E, a 5′ GTP (gray) is added to the sequence to facilitate transcription. (F) Normalized UV melt profiles for M1TH (gray), M1ABsT (orange), M1ET (blue), M1AB (green) from 20 to 85°C in 20 mM HEPES•KOH, pH 7.4, 1 mM MgCl2, 25 mM NaCl and 25 mM KCl. Triplex formation for all constructs is explicitly demonstrated in Supplementary Figure S2. Triplex and duplex melting occur in the same transition for the bimolecular M1AB. All experiments were performed in duplicate. A single representative melting profile is plotted. (G) Triplex melting temperatures (Tm,1) for the unimolecular, bimolecular, and trimolecular constructs, colored as in F. Large changes in Tm,1 for the bi- and tri-molecular RNA relative to the full-length RNA demonstrate the contributions of peripheral elements to triplex stability.
), A-minor interactions (−−), and a wobble base pair (•). A 5′ GTP (grey) is added to the sequence to facilitate transcription. The A-rich tail is colored blue and structural regions are indicated in brackets. (B) The crystal structure of the MALAT1 triple helix core (PDB: 4PLX) (33). The three strands are colored orange, pink, and blue. The two short loops (gray) were engineered to facilitate crystallization. (C) Secondary structure of trimolecular M1ABsT. M1Bs is colored orange, M1A is colored pink, and M1Tail is colored blue, as in B. (D) Secondary structure of bimolecular M1ET, which eliminates the basal linker. The ENE motif is colored cyan and the M1Tail is colored as in C. (E) Secondary structure of bimolecular M1AB, wherein the basal linker is included and the apical P2 helix is truncated. M1A is colored as in C and M1B is colored green. In C–E, a 5′ GTP (gray) is added to the sequence to facilitate transcription. (F) Normalized UV melt profiles for M1TH (gray), M1ABsT (orange), M1ET (blue), M1AB (green) from 20 to 85°C in 20 mM HEPES•KOH, pH 7.4, 1 mM MgCl2, 25 mM NaCl and 25 mM KCl. Triplex formation for all constructs is explicitly demonstrated in Supplementary Figure S2. Triplex and duplex melting occur in the same transition for the bimolecular M1AB. All experiments were performed in duplicate. A single representative melting profile is plotted. (G) Triplex melting temperatures (Tm,1) for the unimolecular, bimolecular, and trimolecular constructs, colored as in F. Large changes in Tm,1 for the bi- and tri-molecular RNA relative to the full-length RNA demonstrate the contributions of peripheral elements to triplex stability.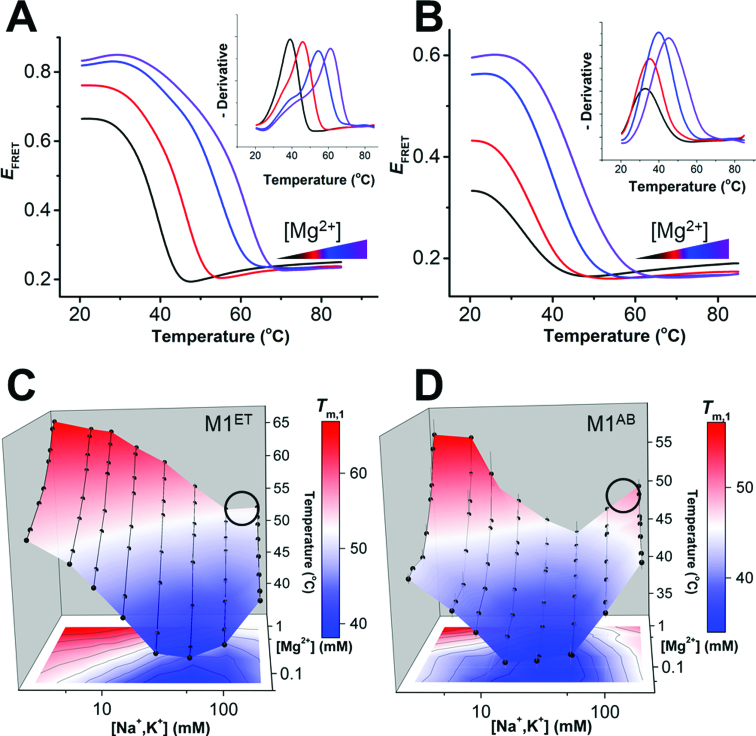
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
