

Review
doi: 10.1016/j.immuni.2018.10.016.
Modulation of Host Immunity by Helminths: The Expanding Repertoire of Parasite Effector Molecules
Affiliations
- PMID: 30462997
- PMCID: PMC6269126
- DOI: 10.1016/j.immuni.2018.10.016
Item in Clipboard
Review
Modulation of Host Immunity by Helminths: The Expanding Repertoire of Parasite Effector Molecules
Immunity.
.
Abstract
Helminths are extraordinarily successful parasites due to their ability to modulate the host immune response. They have evolved a spectrum of immunomodulatory molecules that are now beginning to be defined, heralding a molecular revolution in parasite immunology. These discoveries have the potential both to transform our understanding of parasite adaptation to the host and to develop possible therapies for immune-mediated disease. In this review we will summarize the current state of the art in parasite immunomodulation and discuss perspectives on future areas for research and discovery.
Copyright © 2018 Elsevier Inc. All rights reserved.
Figures
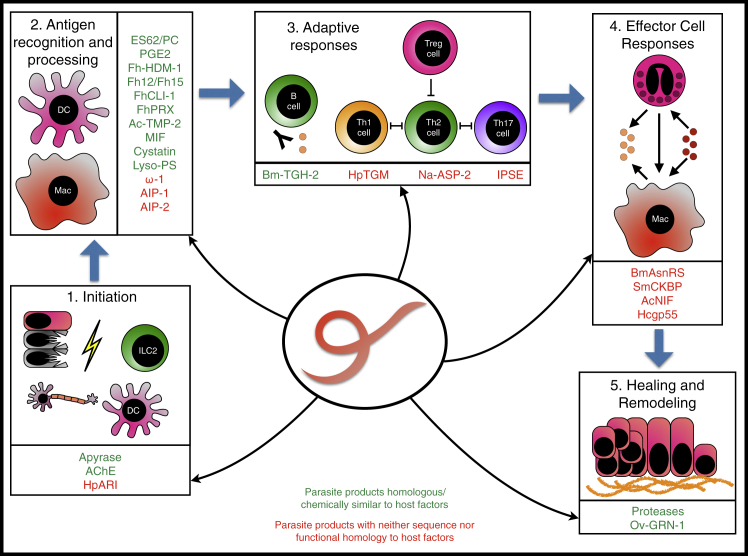
Helminth Modulators Act at All Phases of the Immune Response Modulatory proteins can be homologous to host genes (shared evolutionary ancestry, green) or show no homology (developed de novo, red). The immune response is divided into five phases: (1) initiation, innate recognition of damage and danger; (2) innate cell activation and antigen presentation; (3) adaptive immune responses; (4) effector immune responses; and (5) resolution and healing.
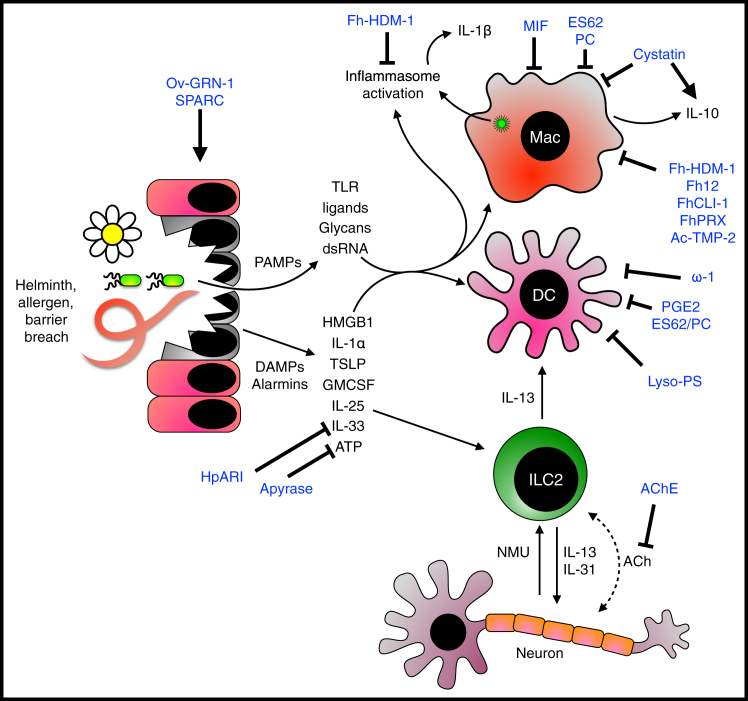
Helminth Modulators that Act on the Early Innate Response to Damage and Danger Helminths and allergens can damage the epithelial layer, resulting in the release of damage associated molecular patterns (DAMPs) and allowing microbial ingress. DAMPs and pathogen associated molecular patterns (PAMPs) can be detected by pattern recognition receptors on dendritic cells and macrophages, resulting in their activation and leading to antigen presentation. Alarmin cytokines such as IL-25, IL-33, and TSLP are also released by epithelial cells and can activate type 2 innate lymphoid cells (ILC2s), which in turn can activate (and be activated by) neurons. ILC2-derived type 2 cytokines aid in the initiation and amplification of the type 2 immune response. Helminth immunomodulators are shown in blue.
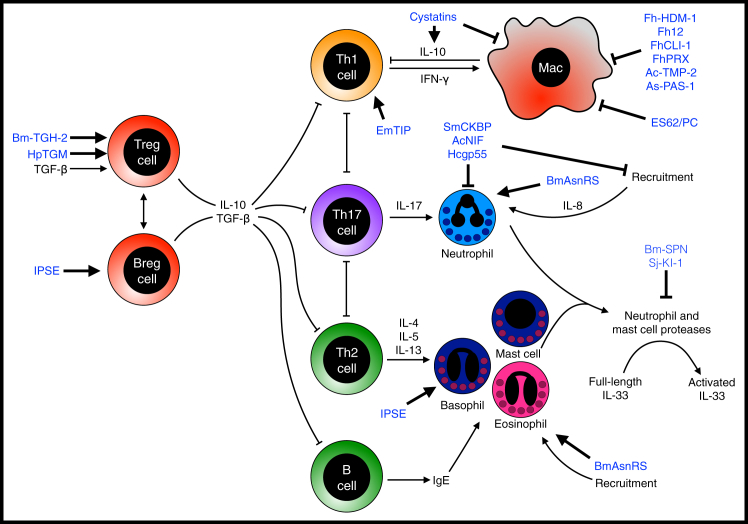
Helminth Modulators that Act on the Adaptive and Effector Immune Responses Immune responses are controlled by helper T cell responses (Th1, Th2, or Th17) and B cell antibody responses. These responses can counter-regulate each other and are suppressed by regulatory T cell or regulatory B cell responses. Effector responses are mediated by T cell-derived cytokines which act on innate effector cells such as macrophages, eosinophils, basophils, and neutrophils. Helminth immunodulators are shown in blue.
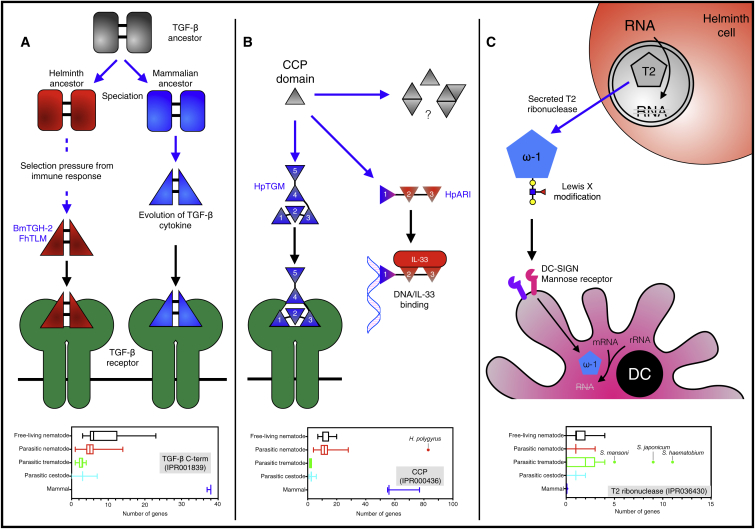
The Evolution and Diversity of Helminth Immunomodulation (A) The TGF-β family is evolutionarily ancient, and nearly all helminth genomes (including free-living helminths) encode TGF-β family members. TGF-β homologs from Brugia malayi (Bm-TGH-2) and Fasciola hepatica (FhTLM) have co-evolved with their host to bind to the mammalian TGF-β receptor. (B) In Heligmosomoides polygyrus, the CCP domain-containing family is over-represented in the genome, and HpTGM (a 5 CCP domain protein) has undergone convergent evolution to bind the TGF-β receptor, despite bearing no homology to the host cytokine. The CCP domain family in H. polygyrus appears highly adaptable, and the IL-33-blocking protein HpARI also consists of 3 CCP domains. (C) T2 ribonucleases are involved in a number of homeostatic processes including RNA recycling, and most helminth genomes contain a member of this family. In the schistosomes, however, the T2 ribonuclease family has undergone expansion, and the T2 ribonucelase omega-1 (ω-1) is secreted into the host, where it gains entry to dendritic cells (DCs) through its Lewis X motifs which are bound by glycan receptors. Once inside the DC, omega-1 degrades host messenger and ribosomal RNA, suppressing DC activation and downstream T cell responses. Blue arrows indicate evolutionary processes, black arrows indicate binding. Graphs show number of members of families in each group of genomes (WormBase ParaSite), represented as box and whiskers representing minimum to maximum values.
References
-
- Akdis M., Akdis C.A. Mechanisms of allergen-specific immunotherapy: multiple suppressor factors at work in immune tolerance to allergens. J. Allergy Clin. Immunol. 2014;133:621–631. - PubMed
-
- Al-Riyami L., Pineda M.A., Rzepecka J., Huggan J.K., Khalaf A.I., Suckling C.J., Scott F.J., Rodgers D.T., Harnett M.M., Harnett W. Designing anti-inflammatory drugs from parasitic worms: a synthetic small molecule analogue of the Acanthocheilonema viteae product ES-62 prevents development of collagen-induced arthritis. J. Med. Chem. 2013;56:9982–10002. - PMC - PubMed
-
- Alvarado R., To J., Lund M.E., Pinar A., Mansell A., Robinson M.W., O’Brien B.A., Dalton J.P., Donnelly S. The immune modulatory peptide FhHDM-1 secreted by the helminth Fasciola hepatica prevents NLRP3 inflammasome activation by inhibiting endolysosomal acidification in macrophages. FASEB J. 2017;31:85–95. - PubMed
-
- Ambadapadi S., Munuswamy-Ramanujam G., Zheng D., Sullivan C., Dai E., Morshed S., McFadden B., Feldman E., Pinard M., McKenna R. Reactive center loop (RCL) peptides derived from serpins display independent coagulation and immune modulating activities. J. Biol. Chem. 2016;291:2874–2887. - PMC - PubMed
-
- Anbu K.A., Joshi P. Identification of a 55 kDa Haemonchus contortus excretory/secretory glycoprotein as a neutrophil inhibitory factor. Parasite Immunol. 2008;30:23–30. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
