

The unusual rainbow trout sex determination gene hijacked the canonical vertebrate gonadal differentiation pathway
- PMID: 30463951
- PMCID: PMC6294932
- DOI: 10.1073/pnas.1803826115
The unusual rainbow trout sex determination gene hijacked the canonical vertebrate gonadal differentiation pathway
Abstract
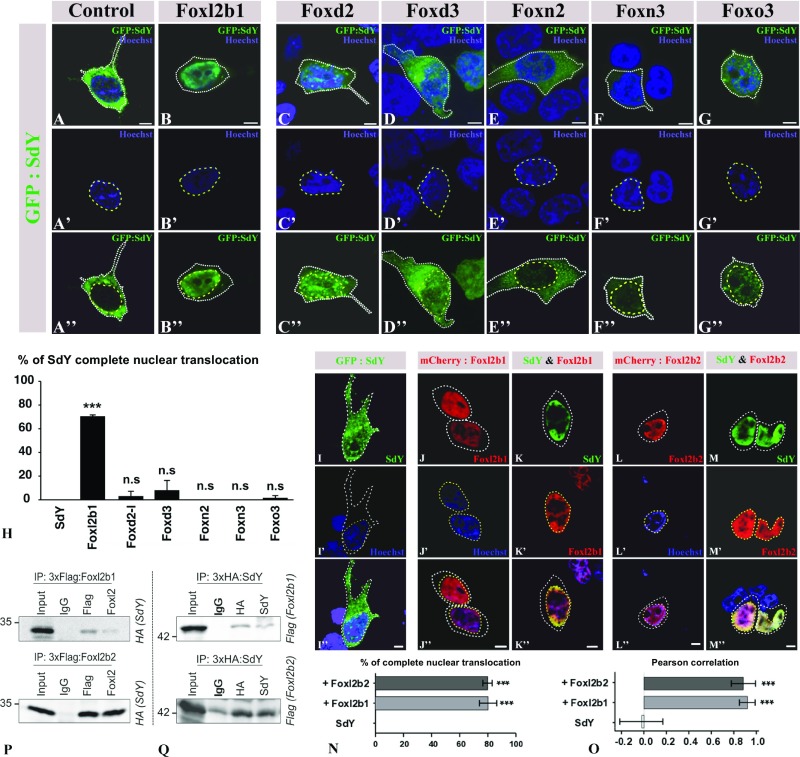
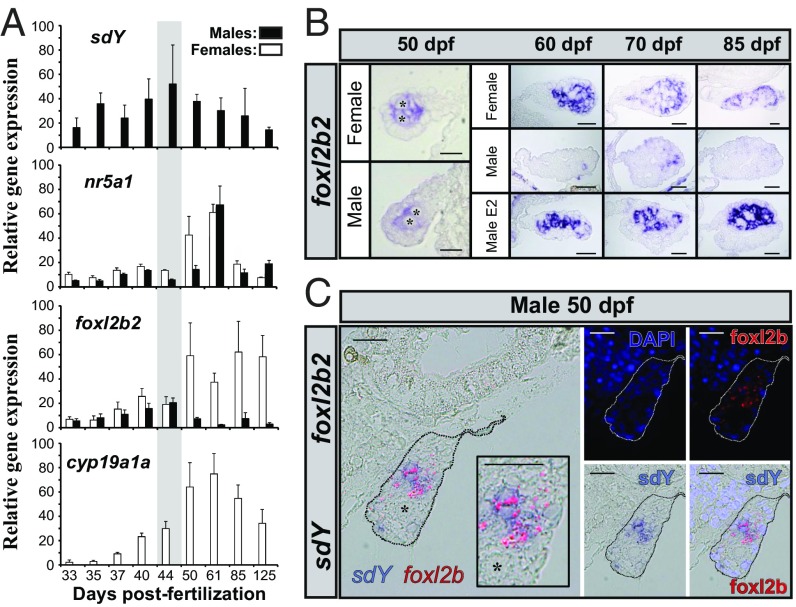
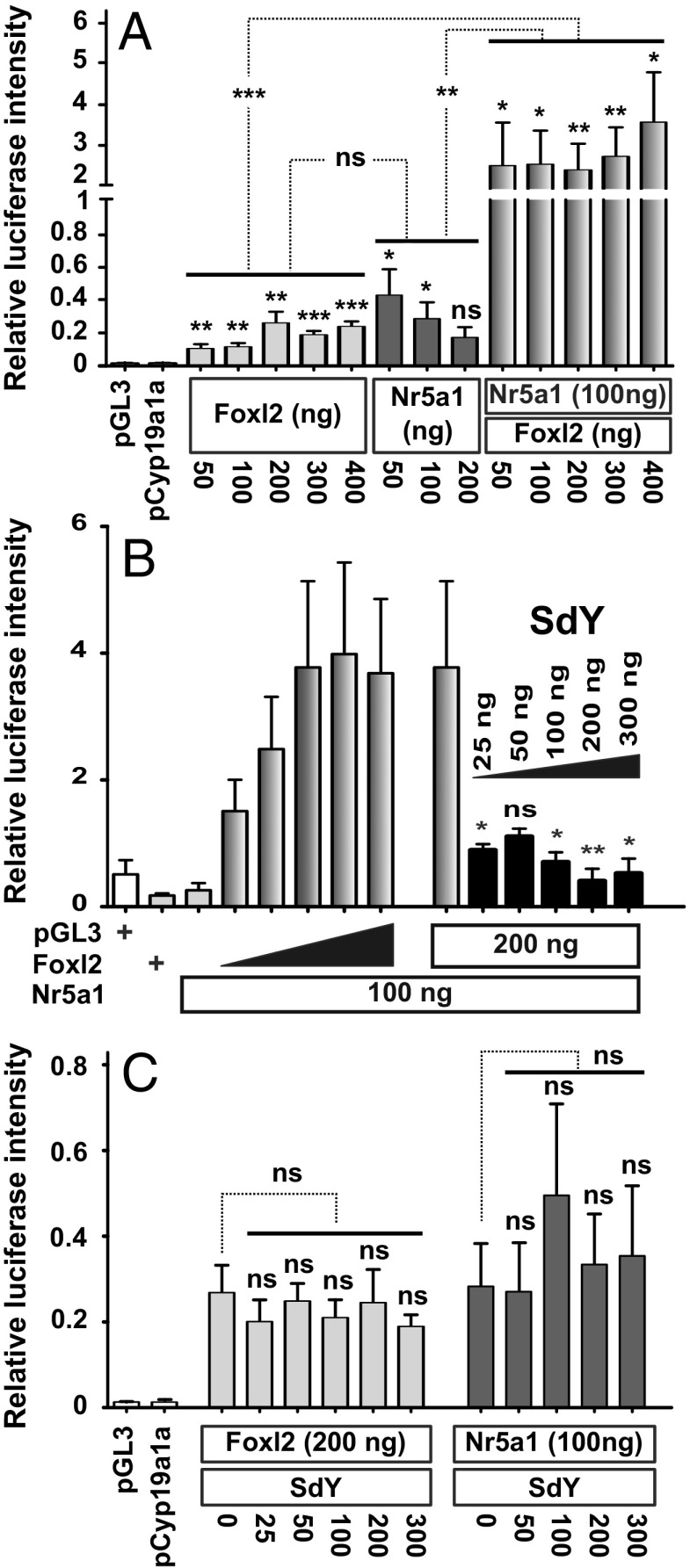
Evolutionary novelties require rewiring of transcriptional networks and/or the evolution of new gene functions. Sex determination (SD), one of the most plastic evolutionary processes, requires such novelties. Studies on the evolution of vertebrate SD revealed that new master SD genes are generally recruited from genes involved in the downstream SD regulatory genetic network. Only a single exception to this rule is currently known in vertebrates: the intriguing case of the salmonid master SD gene (sdY), which arose from duplication of an immune-related gene. This exception immediately posed the question of how a gene outside from the classical sex differentiation cascade could acquire its function as a male SD gene. Here we show that SdY became integrated in the classical vertebrate sex differentiation cascade by interacting with the Forkhead box domain of the female-determining transcription factor, Foxl2. In the presence of Foxl2, SdY is translocated to the nucleus where the SdY:Foxl2 complex prevents activation of the aromatase (cyp19a1a) promoter in cooperation with Nr5a1 (Sf1). Hence, by blocking a positive loop of regulation needed for the synthesis of estrogens in the early differentiating gonad, SdY disrupts a preset female differentiation pathway, consequently allowing testicular differentiation to proceed. These results also suggest that the evolution of unusual vertebrate master sex determination genes recruited from outside the classical pathway like sdY is strongly constrained by their ability to interact with the canonical gonadal differentiation pathway.
Keywords: Forkhead box proteins; evolution; fish; sex determination; sex differentiation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Similar articles
-
An immune-related gene evolved into the master sex-determining gene in rainbow trout, Oncorhynchus mykiss.Curr Biol. 2012 Aug 7;22(15):1423-8. doi: 10.1016/j.cub.2012.05.045. Epub 2012 Jun 21. Curr Biol. 2012. PMID: 22727696
-
A nonfunctional copy of the salmonid sex-determining gene (sdY) is responsible for the "apparent" XY females in Chinook salmon, Oncorhynchus tshawytscha.G3 (Bethesda). 2022 Feb 4;12(2):jkab451. doi: 10.1093/g3journal/jkab451. G3 (Bethesda). 2022. PMID: 35100376 Free PMC article.
-
Roles of forkhead box protein L2 (foxl2) during gonad differentiation and maintenance in a fish, the olive flounder (Paralichthys olivaceus).Reprod Fertil Dev. 2019 Oct;31(11):1742-1752. doi: 10.1071/RD18233. Reprod Fertil Dev. 2019. PMID: 31537253
-
Ovarian aromatase and estrogens: a pivotal role for gonadal sex differentiation and sex change in fish.Gen Comp Endocrinol. 2010 Feb 1;165(3):352-66. doi: 10.1016/j.ygcen.2009.03.002. Epub 2009 Mar 14. Gen Comp Endocrinol. 2010. PMID: 19289125 Review.
-
Just how conserved is vertebrate sex determination?Dev Dyn. 2013 Apr;242(4):380-7. doi: 10.1002/dvdy.23944. Epub 2013 Mar 6. Dev Dyn. 2013. PMID: 23390004 Review.
Cited by
-
The sockeye salmon genome, transcriptome, and analyses identifying population defining regions of the genome.PLoS One. 2020 Oct 29;15(10):e0240935. doi: 10.1371/journal.pone.0240935. eCollection 2020. PLoS One. 2020. PMID: 33119641 Free PMC article.
-
Functional Divergence of Multiple Duplicated Foxl2 Homeologs and Alleles in a Recurrent Polyploid Fish.Mol Biol Evol. 2021 May 4;38(5):1995-2013. doi: 10.1093/molbev/msab002. Mol Biol Evol. 2021. PMID: 33432361 Free PMC article.
-
Insights from a chum salmon (Oncorhynchus keta) genome assembly regarding whole-genome duplication and nucleotide variation influencing gene function.G3 (Bethesda). 2023 Aug 9;13(8):jkad127. doi: 10.1093/g3journal/jkad127. G3 (Bethesda). 2023. PMID: 37293843 Free PMC article.
-
Identification and Genomic Localization of Autosomal sdY Locus in a Population of Atlantic Salmon (Salmo salar).Mar Biotechnol (NY). 2023 Jun;25(3):438-446. doi: 10.1007/s10126-023-10217-4. Epub 2023 May 26. Mar Biotechnol (NY). 2023. PMID: 37233880
-
Screening and Identification of Transcription Factors Potentially Regulating Foxl2 Expression in Chlamys farreri Ovary.Biology (Basel). 2022 Jan 11;11(1):113. doi: 10.3390/biology11010113. Biology (Basel). 2022. PMID: 35053111 Free PMC article.
References
-
- Graves JAM. How to evolve new vertebrate sex determining genes. Dev Dyn. 2013;242:354–359. - PubMed
-
- Windley SP, Wilhelm D. Signaling pathways involved in mammalian sex determination and gonad development. Sex Dev. 2015;9:297–315. - PubMed
-
- Kikuchi K, Hamaguchi S. Novel sex-determining genes in fish and sex chromosome evolution. Dev Dyn. 2013;242:339–353. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
