

TDP-43 and RNA form amyloid-like myo-granules in regenerating muscle
- PMID: 30464263
- PMCID: PMC6324568
- DOI: 10.1038/s41586-018-0665-2
TDP-43 and RNA form amyloid-like myo-granules in regenerating muscle
Abstract
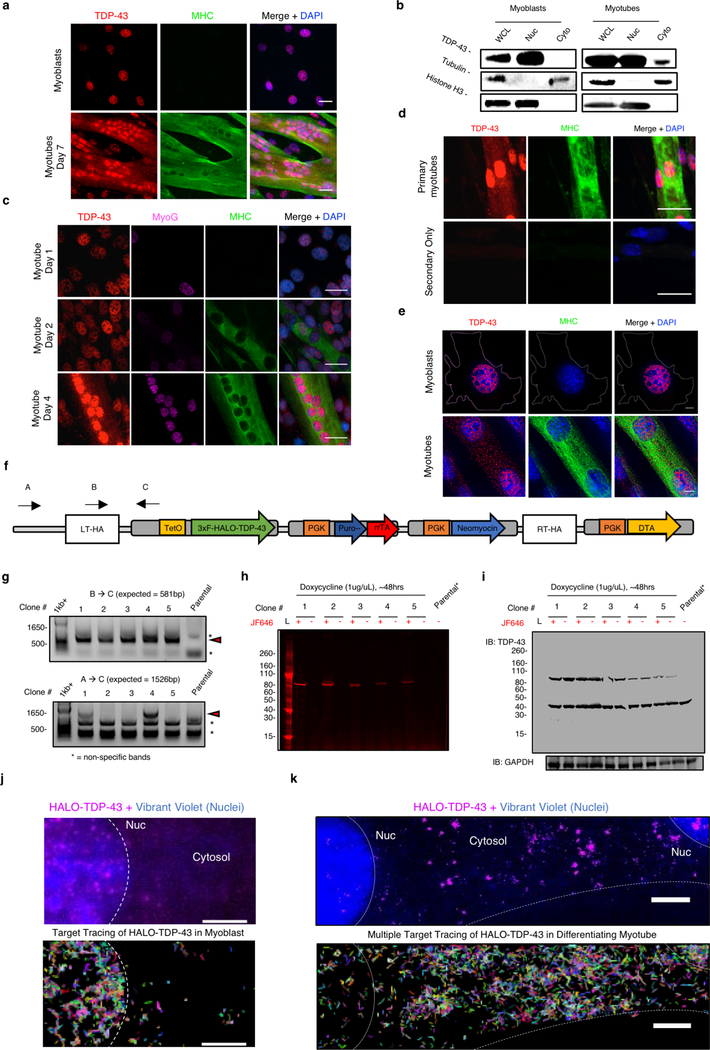
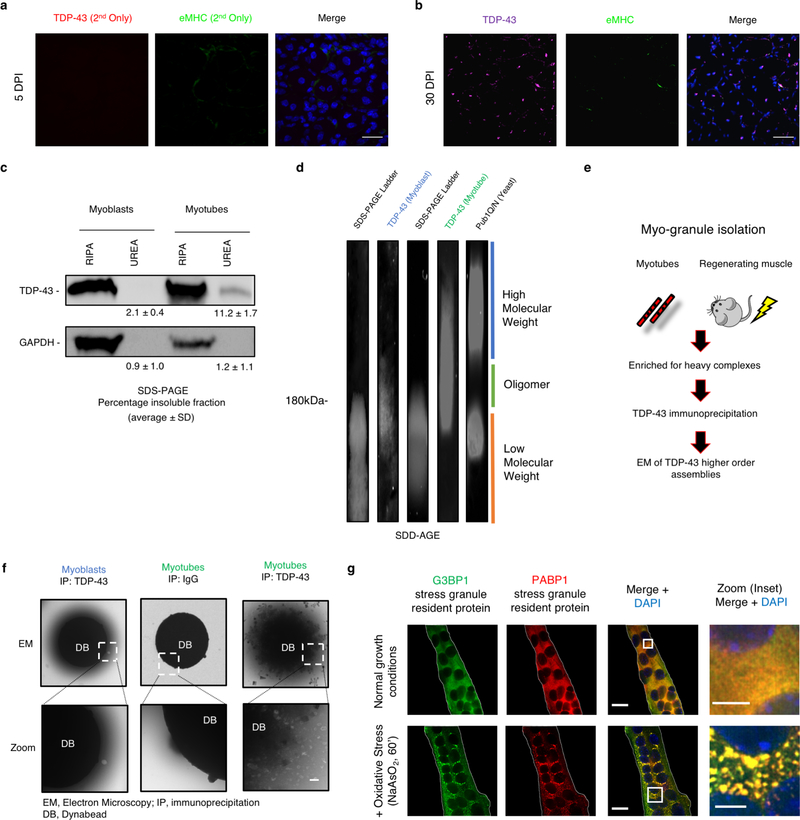
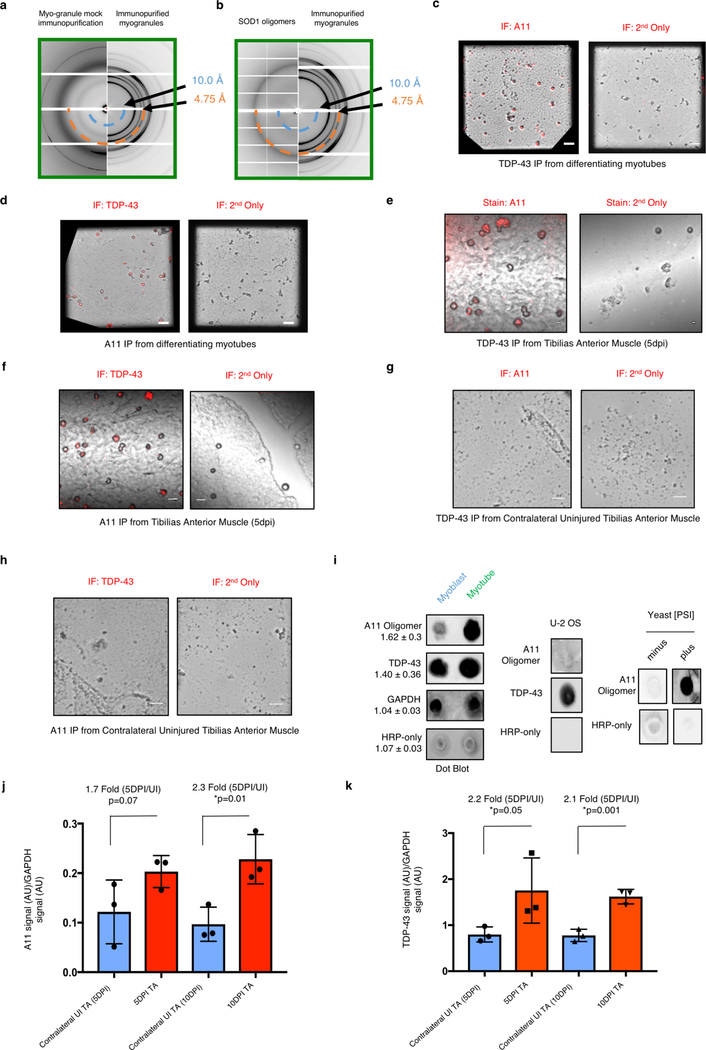
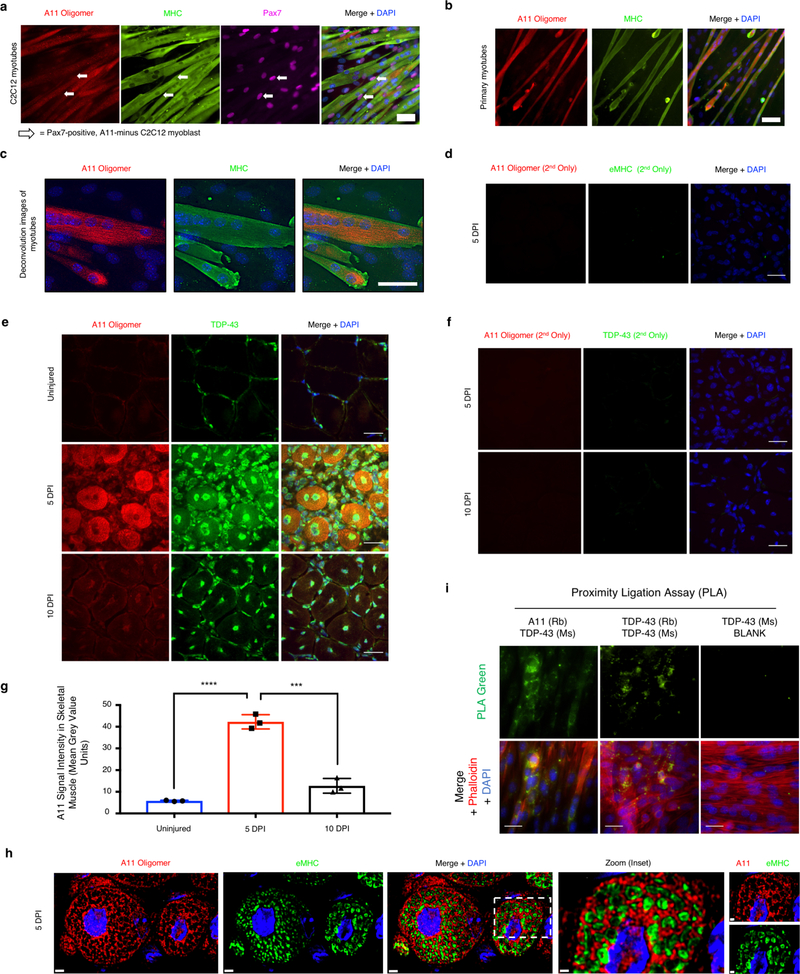
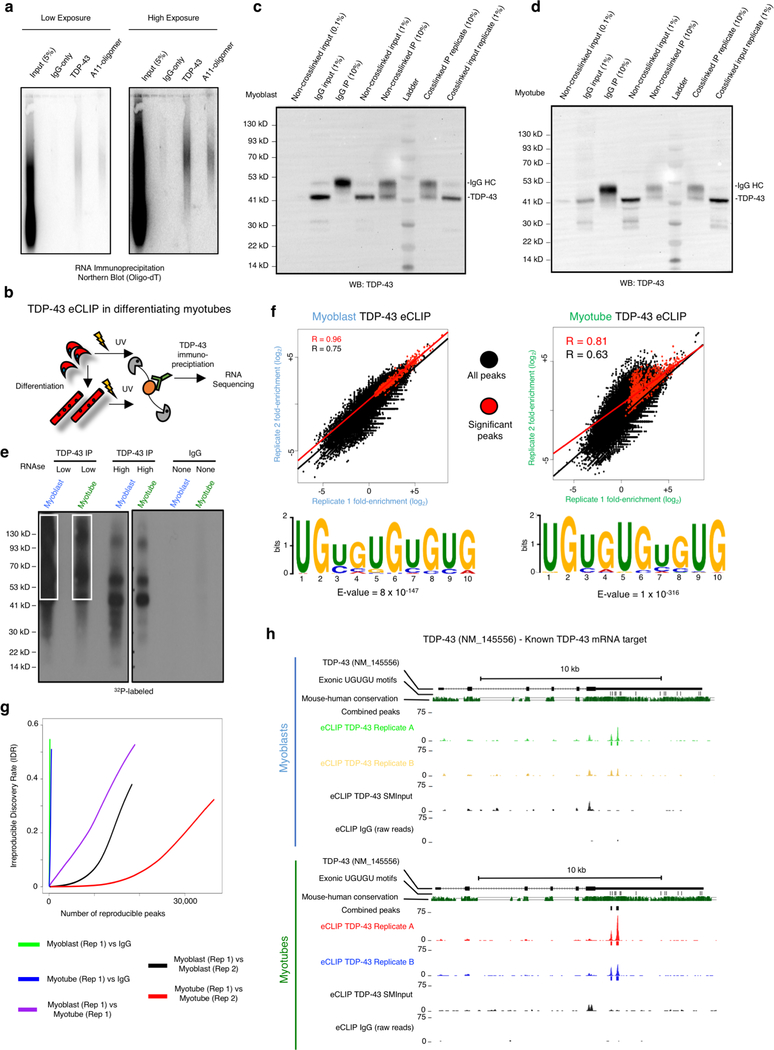
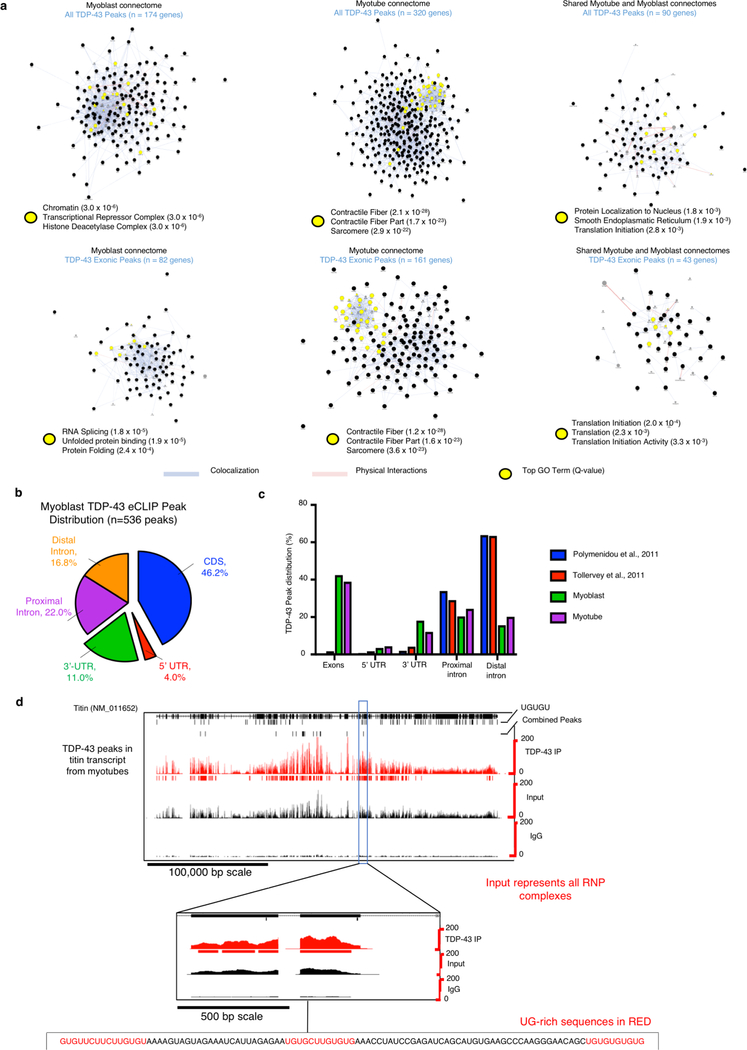
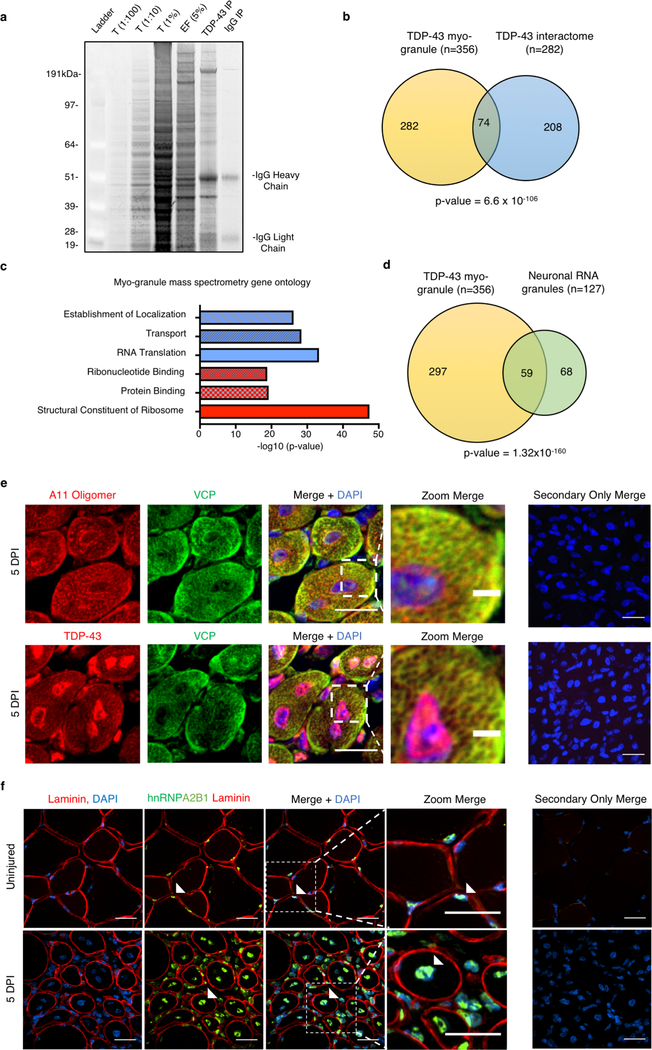
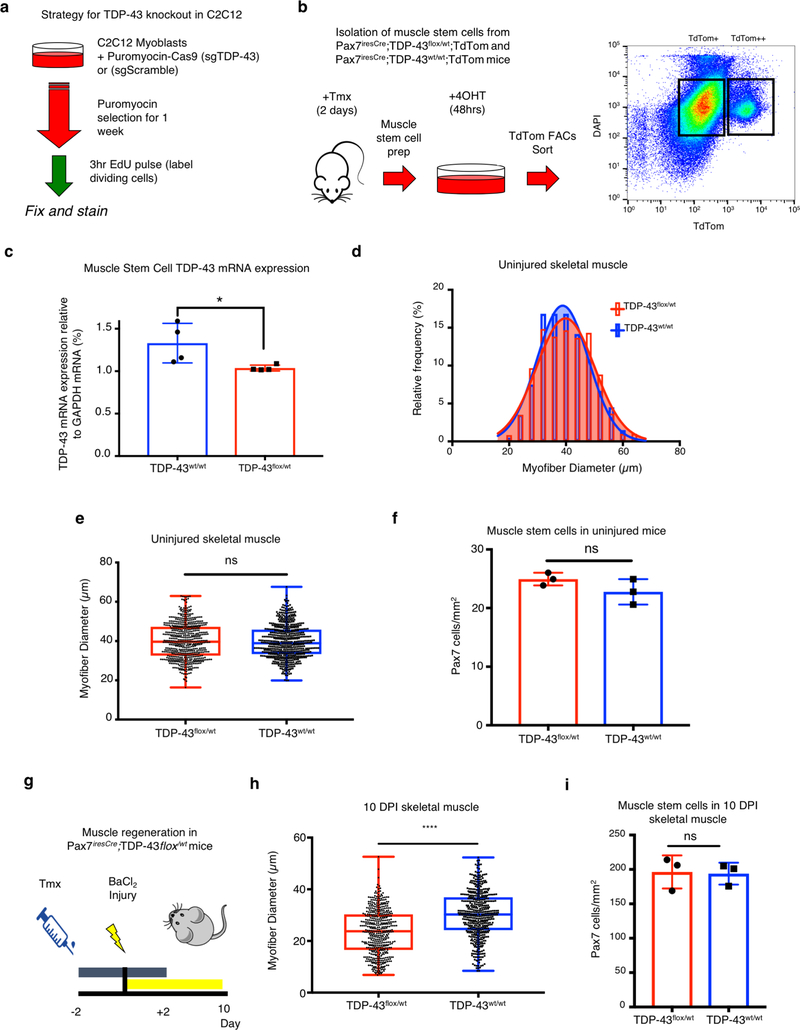
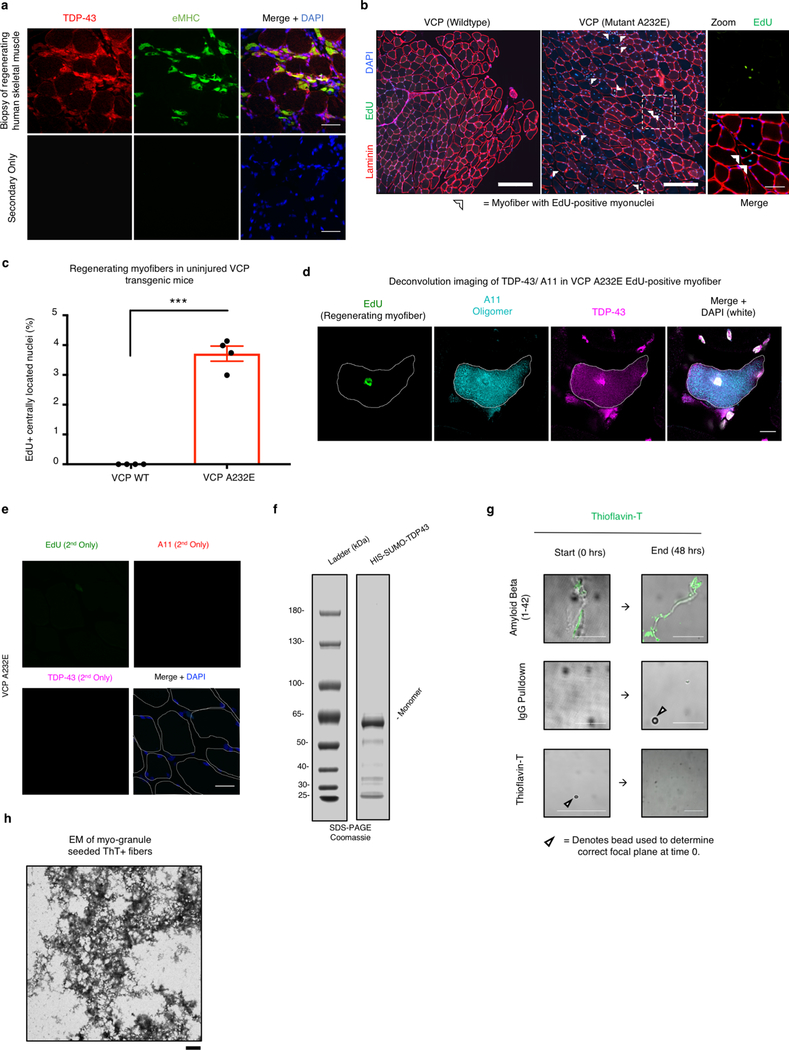
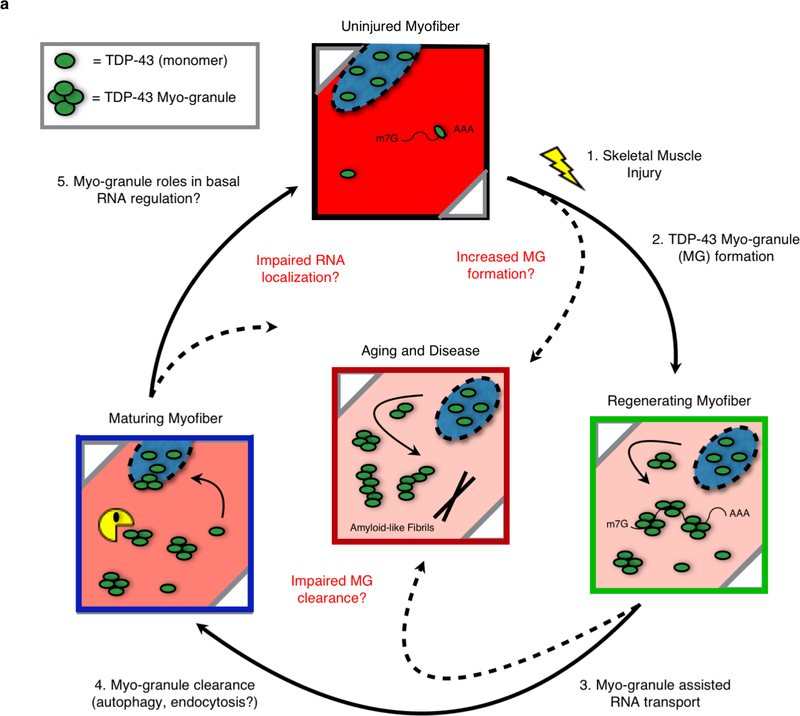
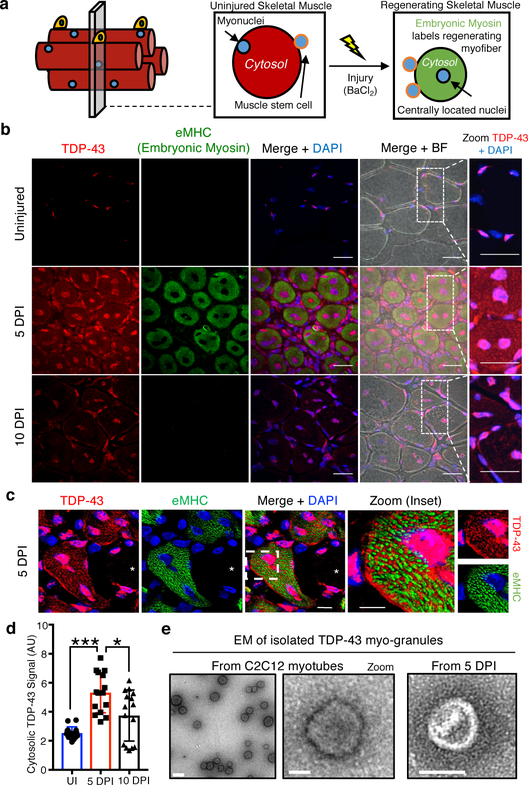
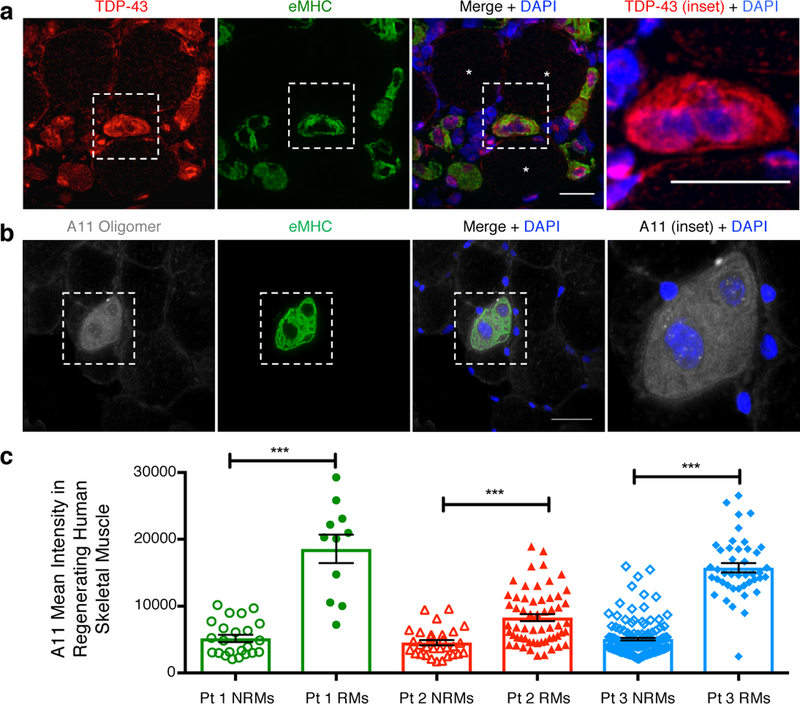
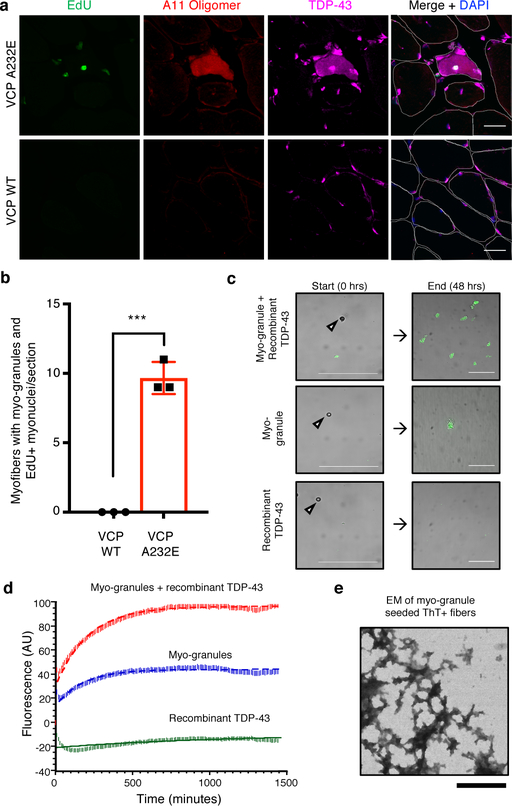
A dominant histopathological feature in neuromuscular diseases, including amyotrophic lateral sclerosis and inclusion body myopathy, is cytoplasmic aggregation of the RNA-binding protein TDP-43. Although rare mutations in TARDBP-the gene that encodes TDP-43-that lead to protein misfolding often cause protein aggregation, most patients do not have any mutations in TARDBP. Therefore, aggregates of wild-type TDP-43 arise in most patients by an unknown mechanism. Here we show that TDP-43 is an essential protein for normal skeletal muscle formation that unexpectedly forms cytoplasmic, amyloid-like oligomeric assemblies, which we call myo-granules, during regeneration of skeletal muscle in mice and humans. Myo-granules bind to mRNAs that encode sarcomeric proteins and are cleared as myofibres mature. Although myo-granules occur during normal skeletal-muscle regeneration, myo-granules can seed TDP-43 amyloid fibrils in vitro and are increased in a mouse model of inclusion body myopathy. Therefore, increased assembly or decreased clearance of functionally normal myo-granules could be the source of cytoplasmic TDP-43 aggregates that commonly occur in neuromuscular disease.
Conflict of interest statement
Author Information
The authors declare no competing interests.
Figures


Comment in
-
A neurodegenerative-disease protein forms beneficial aggregates in healthy muscle.Nature. 2018 Nov;563(7732):477-478. doi: 10.1038/d41586-018-07141-2. Nature. 2018. PMID: 30459367 No abstract available.
-
TDP-43 in the muscles: friend or foe?Nat Rev Rheumatol. 2019 Jan;15(1):1. doi: 10.1038/s41584-018-0138-x. Nat Rev Rheumatol. 2019. PMID: 30531851 No abstract available.
-
Myo-granules Connect Physiology and Pathophysiology.J Exp Neurosci. 2019 Apr 12;13:1179069519842157. doi: 10.1177/1179069519842157. eCollection 2019. J Exp Neurosci. 2019. PMID: 31019368 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
- F30 AR068881/AR/NIAMS NIH HHS/United States
- R01 AR070360/AR/NIAMS NIH HHS/United States
- R01 GM045443/GM/NIGMS NIH HHS/United States
- R35 GM119575/GM/NIGMS NIH HHS/United States
- T32 GM008497/GM/NIGMS NIH HHS/United States
- P30 CA046934/CA/NCI NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- F30 NS093682/NS/NINDS NIH HHS/United States
- R37 GM045443/GM/NIGMS NIH HHS/United States
- R01 GM099836/GM/NIGMS NIH HHS/United States
- T32 GM008752/GM/NIGMS NIH HHS/United States
- R01 AR049446/AR/NIAMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
