

From solution to surface to filament: actin flux into branched networks
- PMID: 30470968
- PMCID: PMC6297094
- DOI: 10.1007/s12551-018-0469-5
From solution to surface to filament: actin flux into branched networks
Abstract
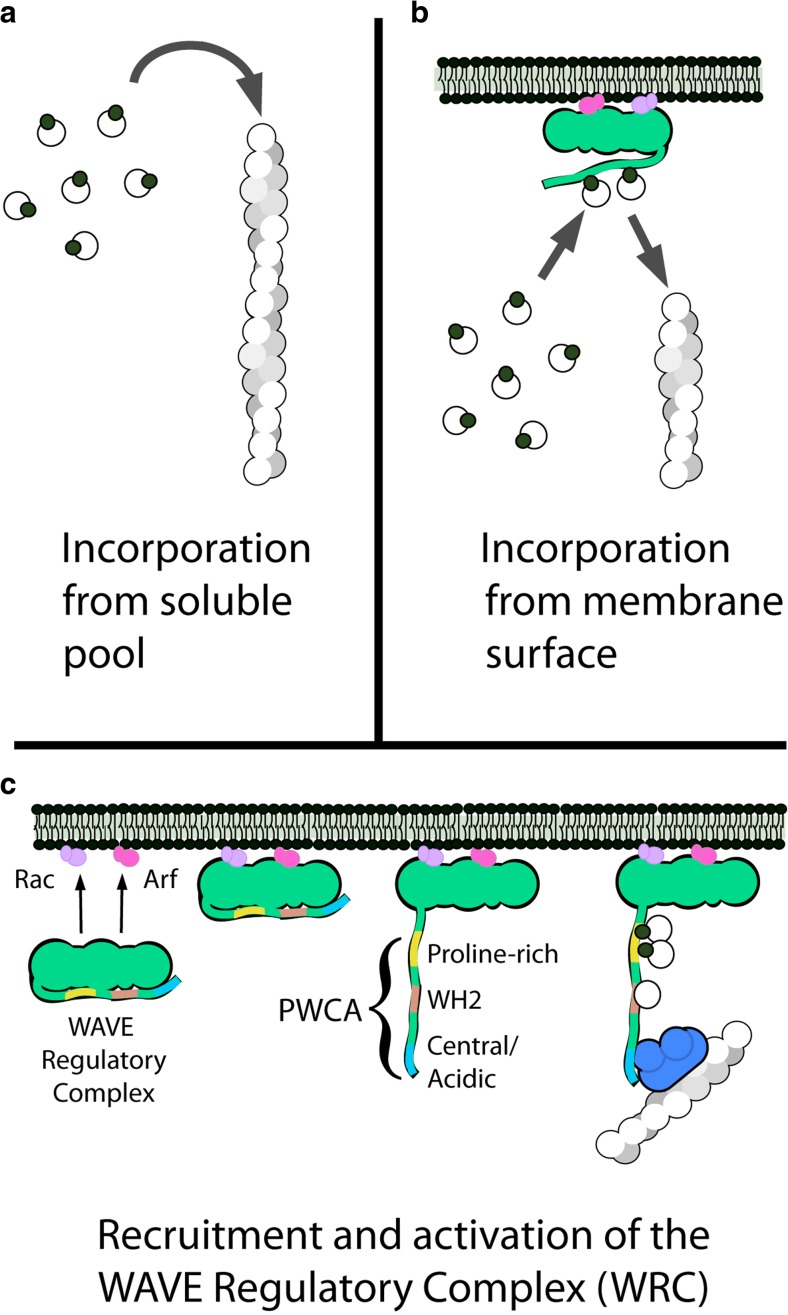
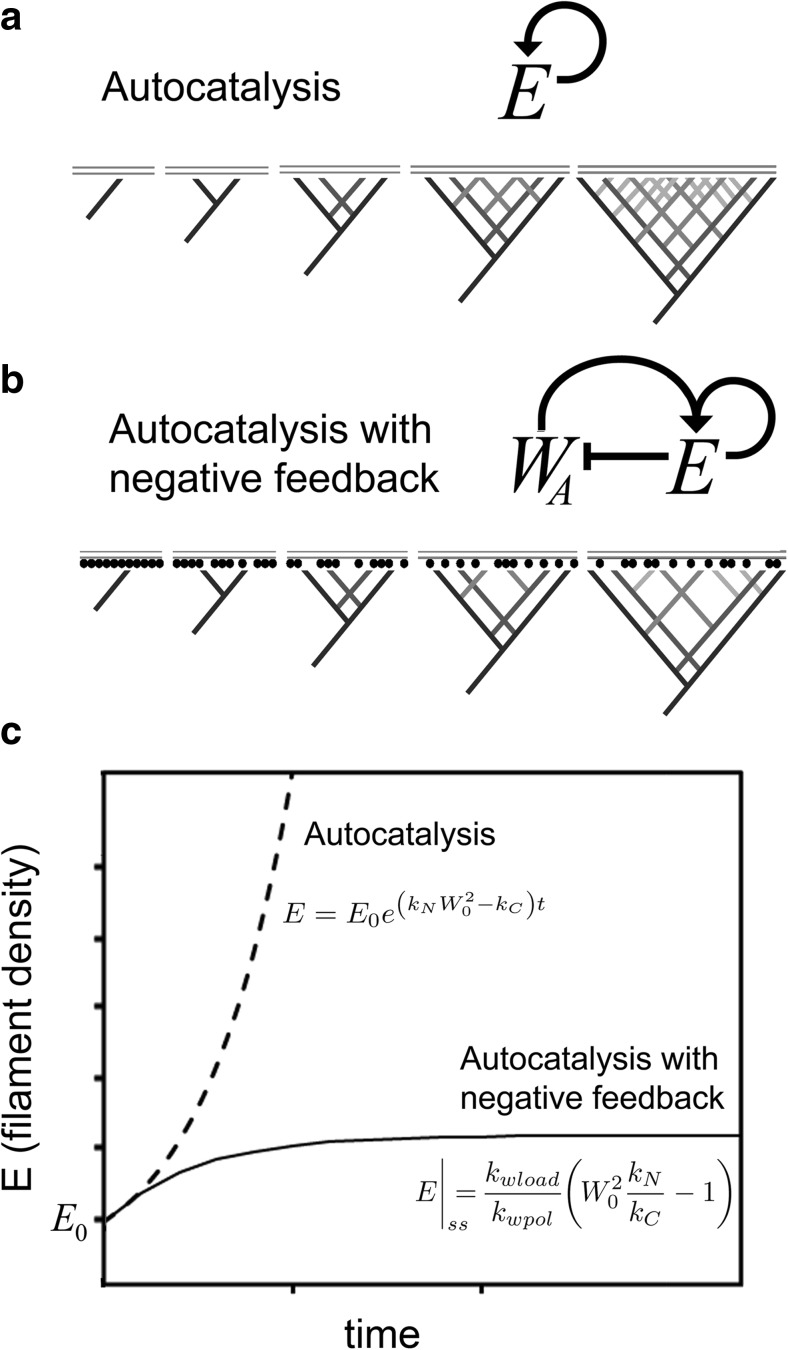
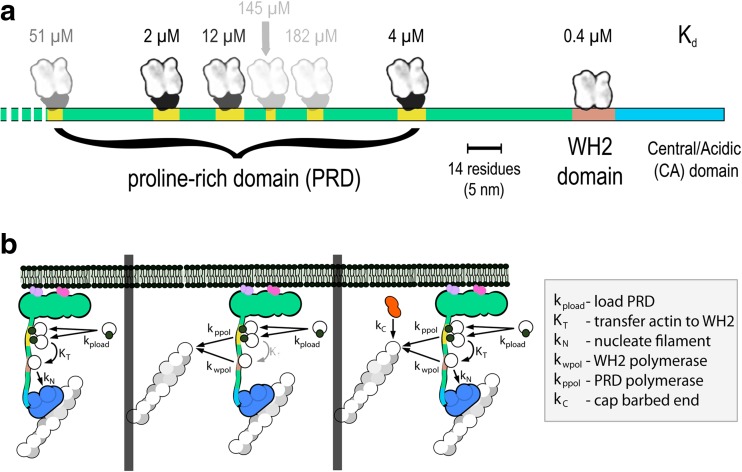
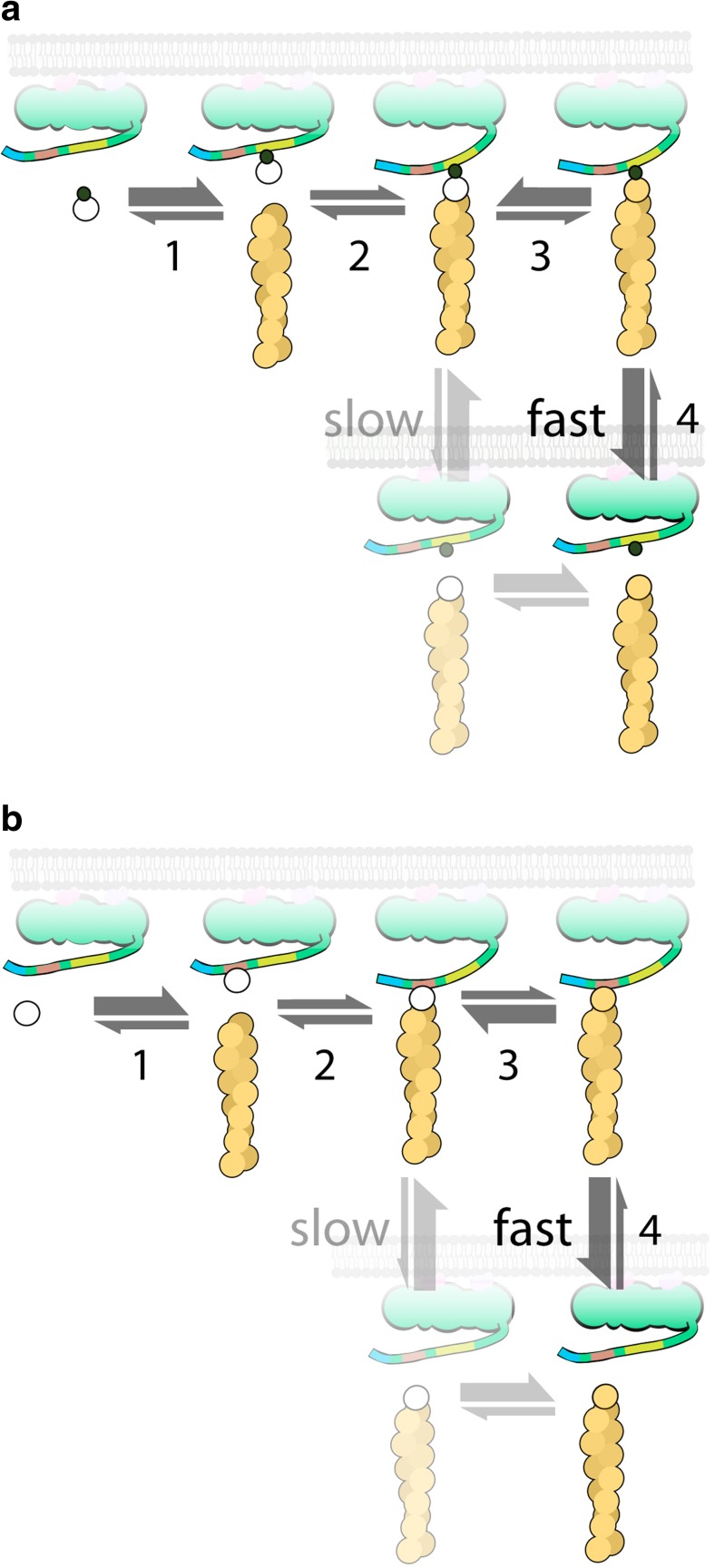
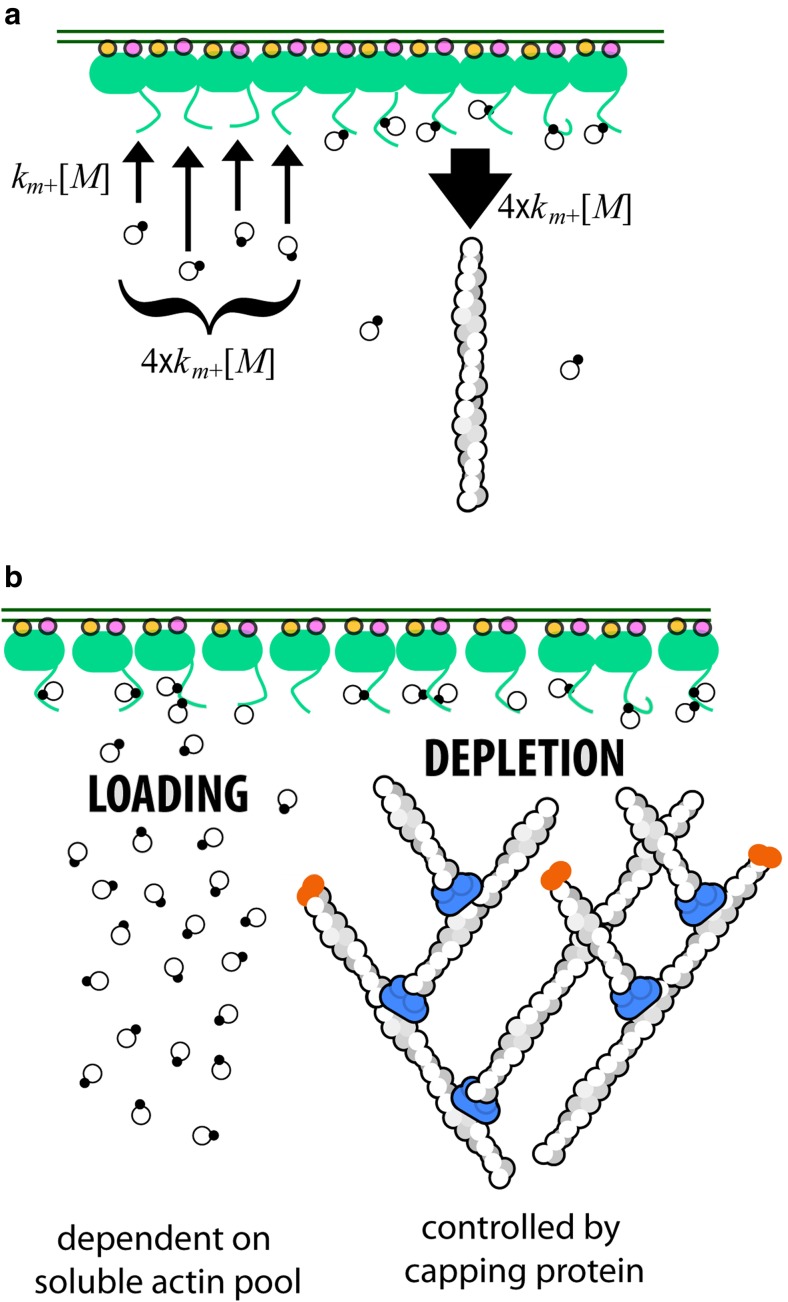
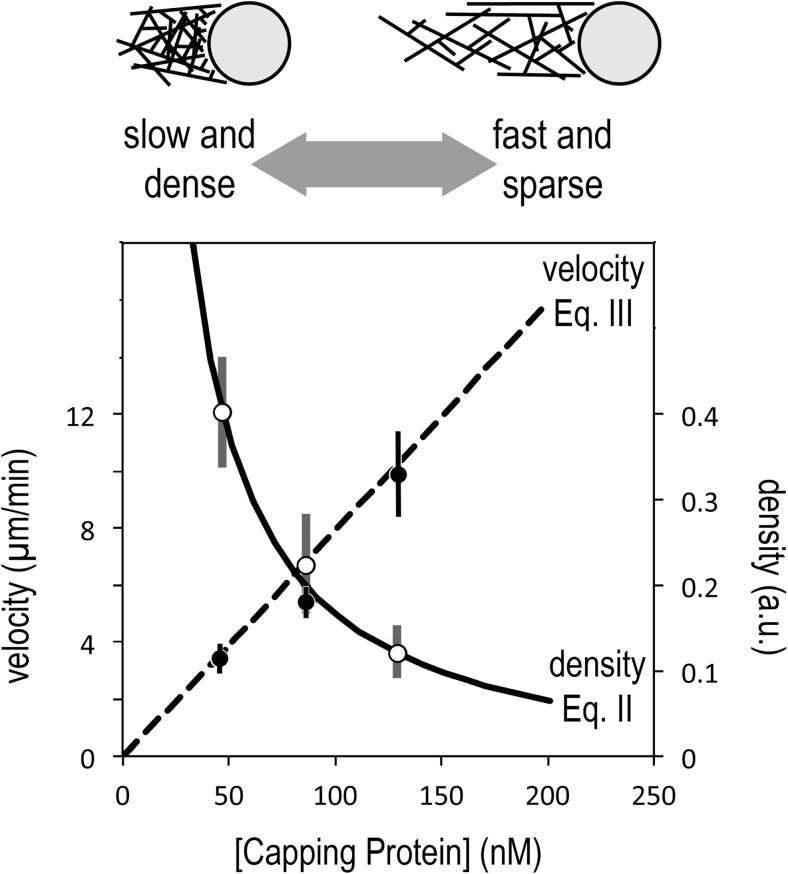
The actin cytoskeleton comprises a set of filament networks that perform essential functions in eukaryotic cells. The idea that actin filaments incorporate monomers directly from solution forms both the "textbook picture" of filament elongation and a conventional starting point for quantitative modeling of cellular actin dynamics. Recent work, however, reveals that filaments created by two major regulators, the formins and the Arp2/3 complex, incorporate monomers delivered by nearby proteins. Specifically, actin enters Arp2/3-generated networks via binding sites on nucleation-promoting factors clustered on membrane surfaces. Here, we describe three functions of this surface-associated actin monomer pool: (1) regulating network density via product inhibition of the Arp2/3 complex, (2) accelerating filament elongation as a distributive polymerase, and (3) converting profilin-actin into a substrate for the Arp2/3 complex. These linked functions control the architecture of branched networks and explain how capping protein enhances their growth.
Keywords: Actin cytoskeleton; Actin filament; Arp2/3 complex; Branched actin network; Capping protein; WASP family protein.
Conflict of interest statement
Conflict of interest
R. Dyche Mullins declares that he has no conflict of interest. Peter Bieling declares that he has no conflict of interest. Daniel Fletcher declares that he has no conflict of interest.
Ethical approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Figures
References
-
- Achard V, Martiel JL, Michelot A, Guérin C, Reymann AC, Blanchoin L, Boujemaa-Paterski R. A "primer"-based mechanism underlies branched actin filament network formation and motility. Curr Biol. 2010;20(5):423–428. - PubMed
-
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P. Molecular biology of the cell. 6. New York: Garland Science; 2015.
-
- Bernheim-Groswasser A, Wiesner S, Golsteyn RM, Carlier MF, Sykes C. The dynamics of actin-based motility depend on surface parameters. Nature. 2002;417(6886):308–311. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
