

Remodeling of cholinergic input to the hippocampus after noise exposure and tinnitus induction in Guinea pigs
- PMID: 30471164
- PMCID: PMC7357289
- DOI: 10.1002/hipo.23058
Remodeling of cholinergic input to the hippocampus after noise exposure and tinnitus induction in Guinea pigs
Abstract
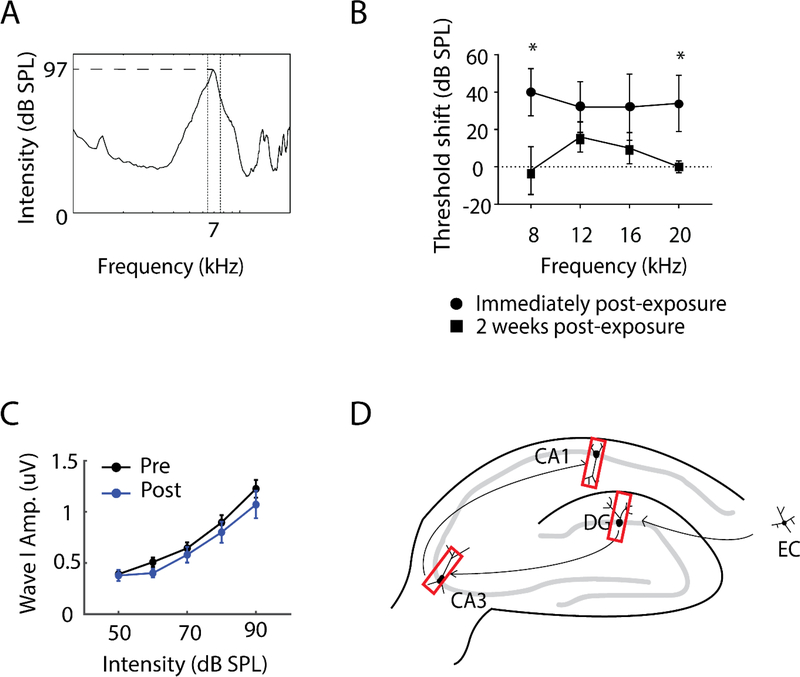
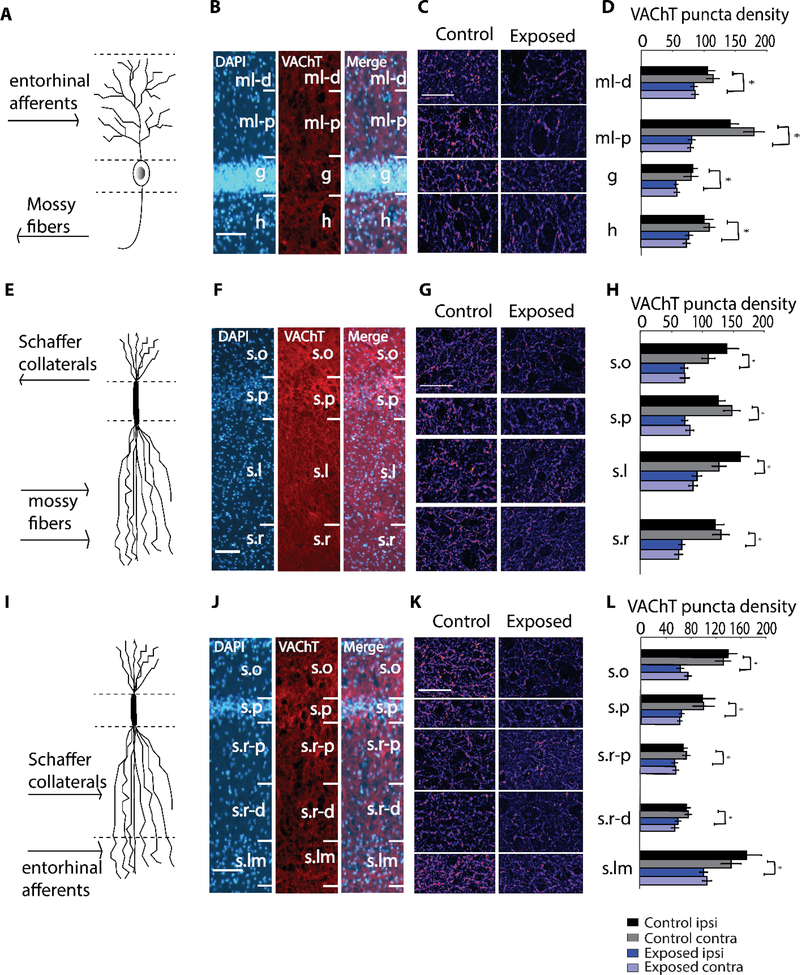
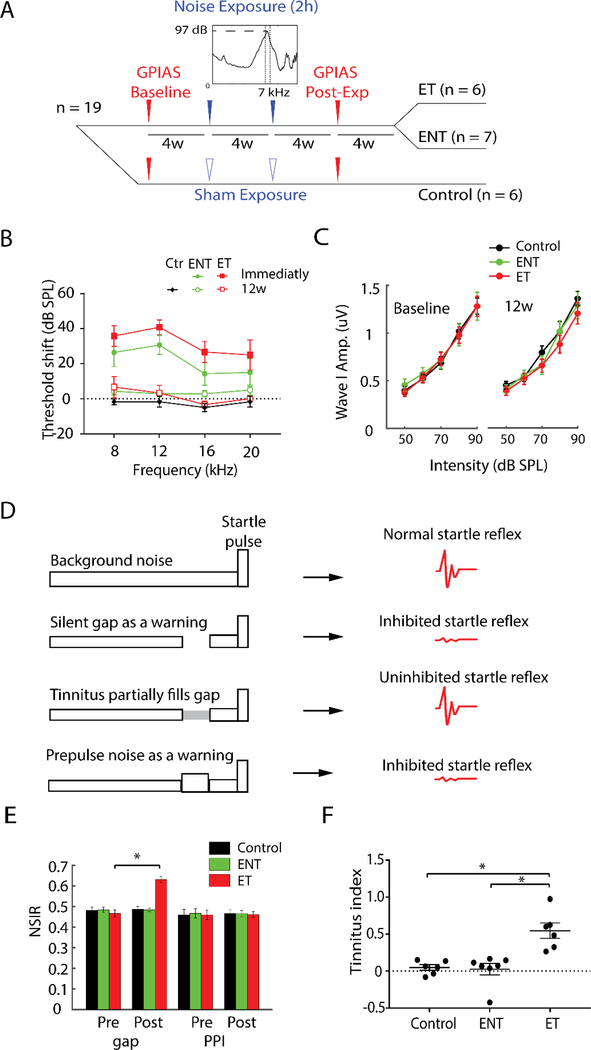
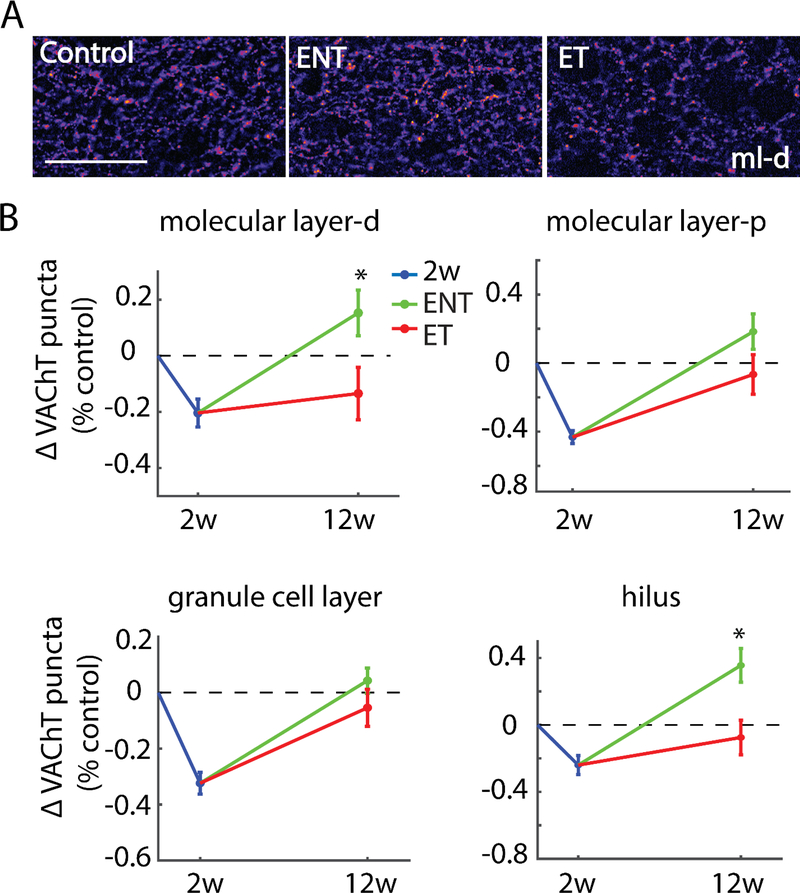
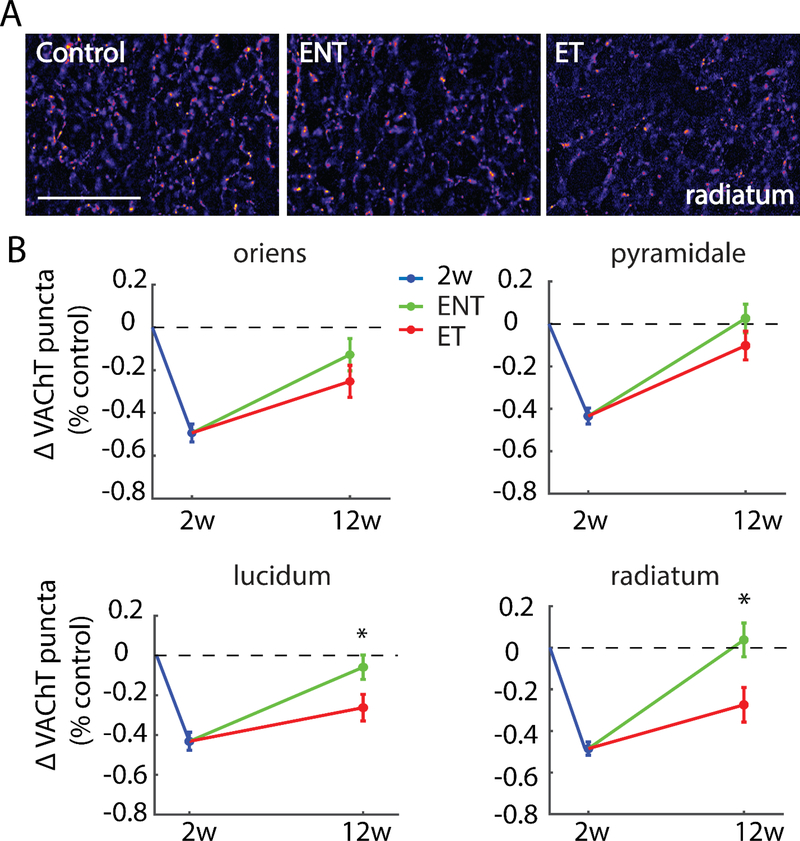
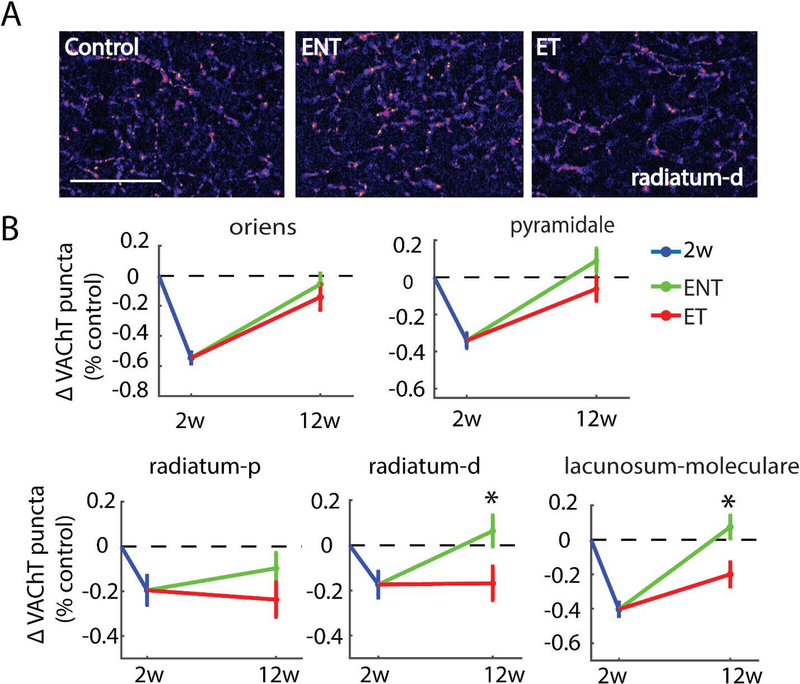
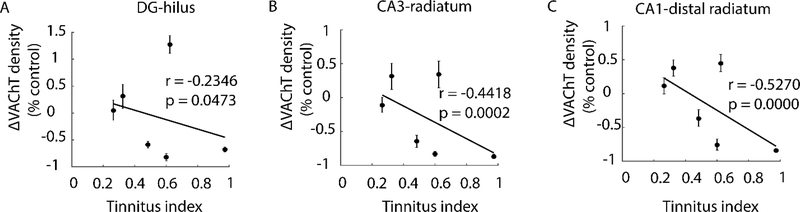
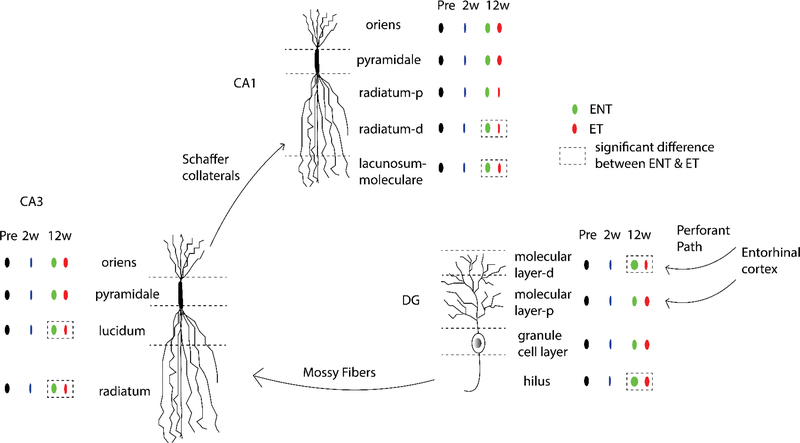
Here, we investigate remodeling of hippocampal cholinergic inputs after noise exposure and determine the relevance of these changes to tinnitus. To assess the effects of noise exposure on the hippocampus, guinea pigs were exposed to unilateral noise for 2 hr and 2 weeks later, immunohistochemistry was performed on hippocampal sections to examine vesicular acetylcholine transporter (VAChT) expression. To evaluate whether the changes in VAChT were relevant to tinnitus, another group of animals was exposed to the same noise band twice to induce tinnitus, which was assessed using gap-prepulse Inhibition of the acoustic startle (GPIAS) 12 weeks after the first noise exposure, followed by immunohistochemistry. Acoustic Brainstem Response (ABR) thresholds were elevated immediately after noise exposure for all experimental animals but returned to baseline levels several days after noise exposure. ABR wave I amplitude-intensity functions did not show any changes after 2 or 12 weeks of recovery compared to baseline levels. In animals assessed 2-weeks following noise-exposure, hippocampal VAChT puncta density decreased on both sides of the brain by 20-60% in exposed animals. By 12 weeks following the initial noise exposure, changes in VAChT puncta density largely recovered to baseline levels in exposed animals that did not develop tinnitus, but remained diminished in animals that developed tinnitus. These tinnitus-specific changes were particularly prominent in hippocampal synapse-rich layers of the dentate gyrus and areas CA3 and CA1, and VAChT density in these regions negatively correlated with tinnitus severity. The robust changes in VAChT labeling in the hippocampus 2 weeks after noise exposure suggest involvement of this circuitry in auditory processing. After chronic tinnitus induction, tinnitus-specific changes occurred in synapse-rich layers of the hippocampus, suggesting that synaptic processing in the hippocampus may play an important role in the pathophysiology of tinnitus.
Keywords: auditory; limbic system; memory; neuroplasticity; vesicular acetylcholine transporter (VAChT).
© 2018 Wiley Periodicals, Inc.
Figures
References
-
- Altman NS (1992). An introduction to kernel and nearest-neighbor nonparametric regression. The American Statistician, 46(3), 10. doi: 10.1080/00031305.1992.10475879 - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
