

Mutations in Recessive Congenital Ichthyoses Illuminate the Origin and Functions of the Corneocyte Lipid Envelope
- PMID: 30471252
- PMCID: PMC11249047
- DOI: 10.1016/j.jid.2018.11.005
Mutations in Recessive Congenital Ichthyoses Illuminate the Origin and Functions of the Corneocyte Lipid Envelope
Abstract
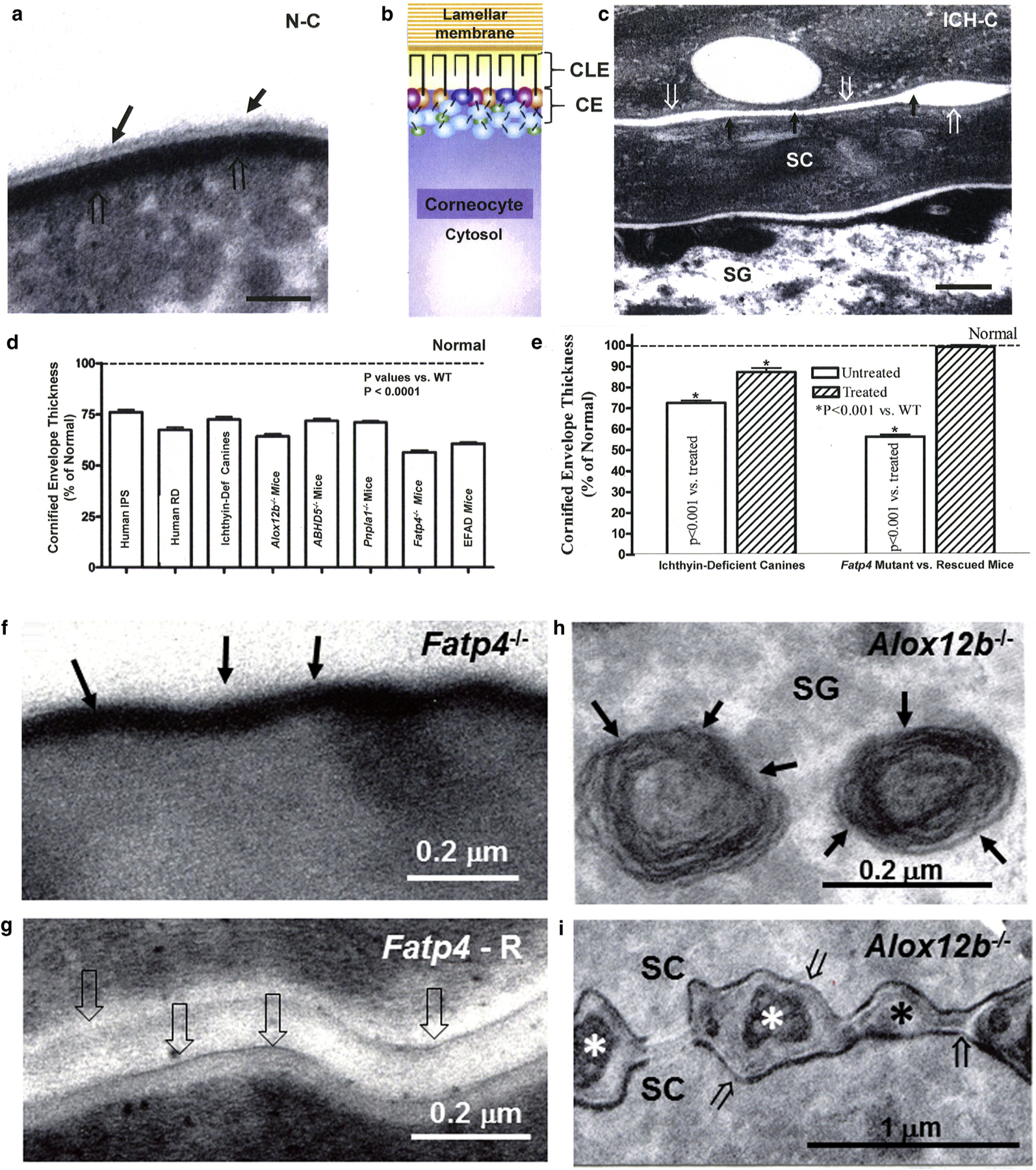
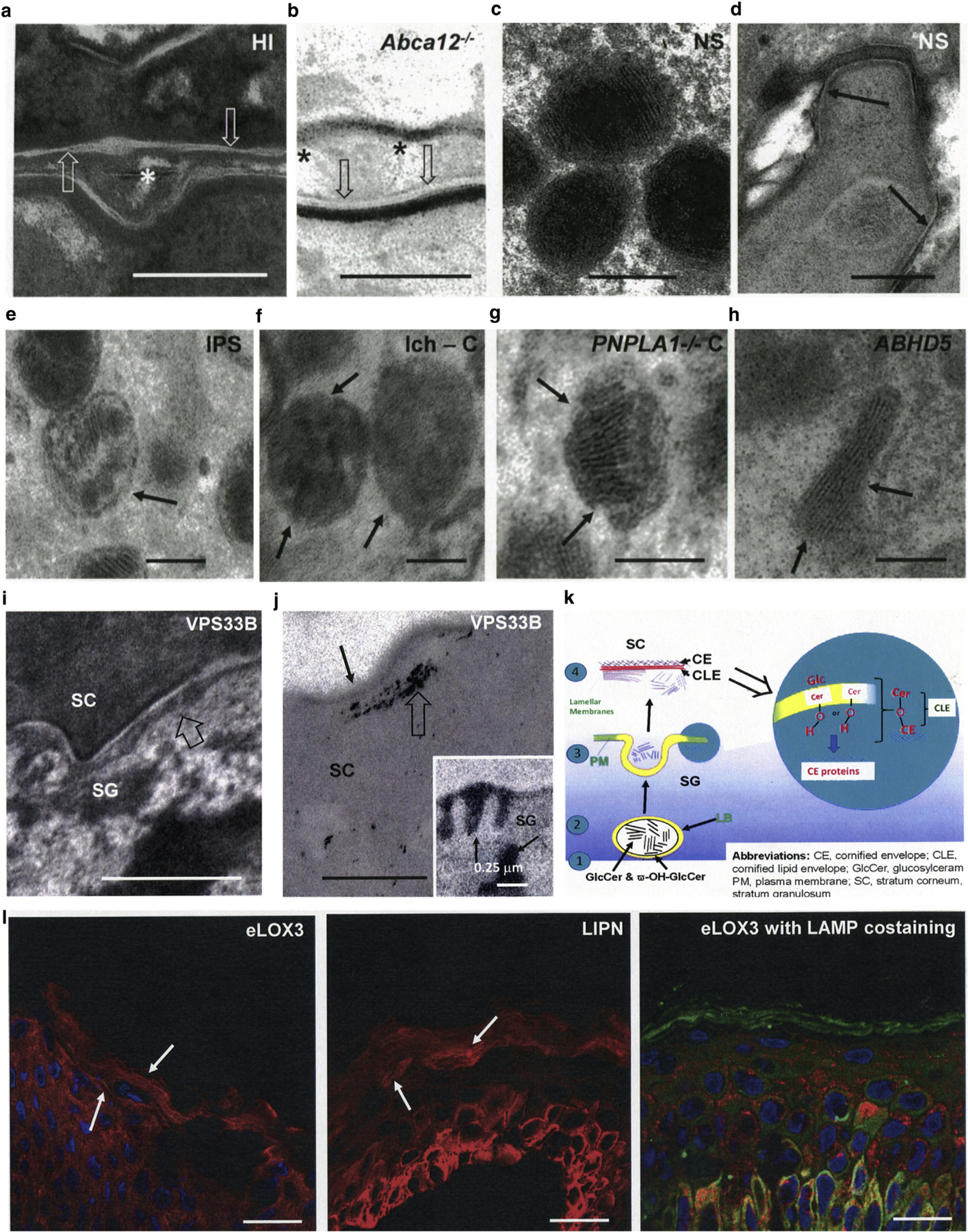
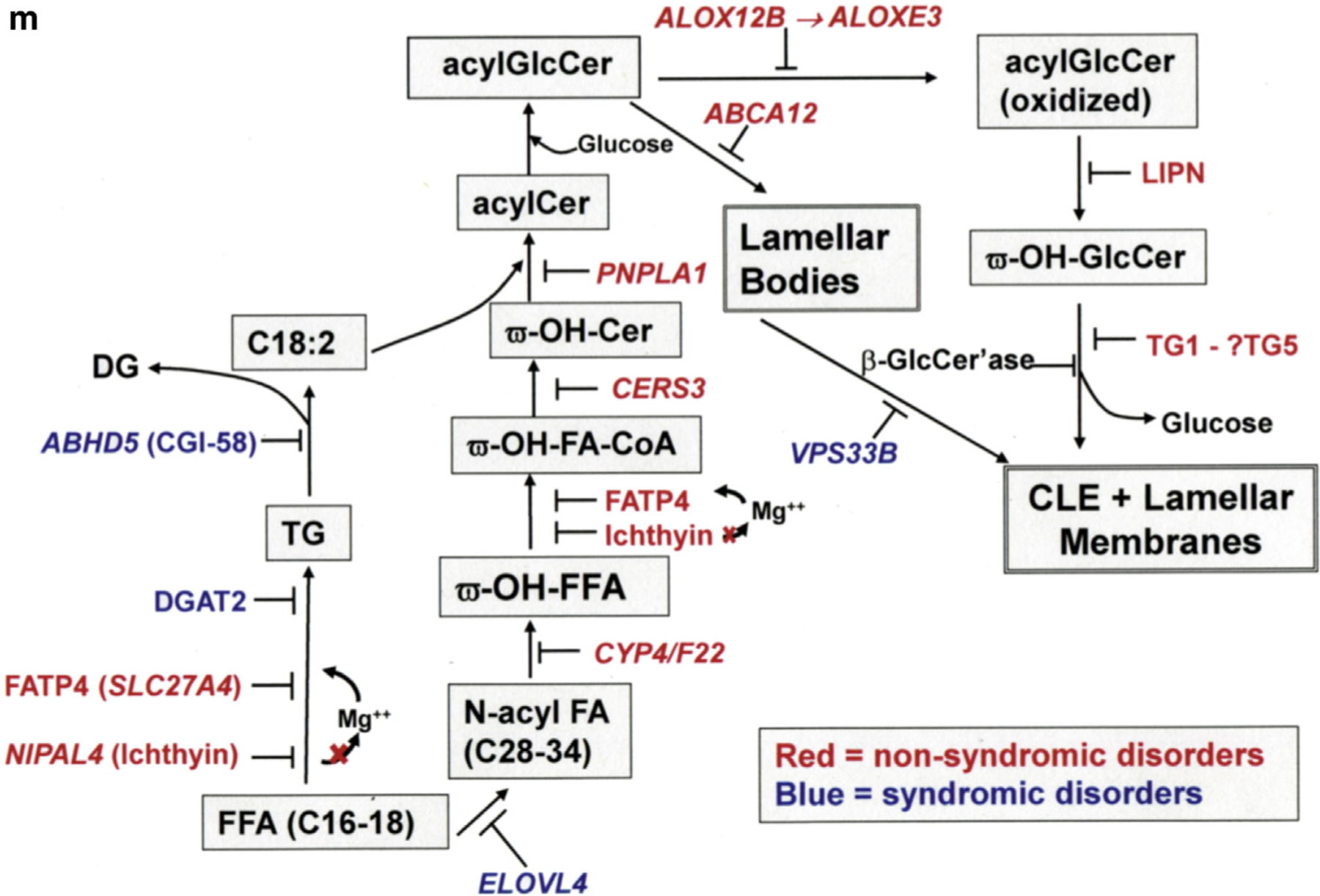
The corneocyte lipid envelope (CLE), a monolayer of ω-hydroxyceramides whose function(s) remain(s) uncertain, is absent in patients with autosomal recessive congenital ichthyoses with mutations in enzymes that regulate epidermal lipid synthesis. Secreted lipids fail to transform into lamellar membranes in certain autosomal recessive congenital ichthyosis epidermis, suggesting the CLE provides a scaffold for the extracellular lamellae. However, because cornified envelopes are attenuated in these autosomal recessive congenital ichthyoses, the CLE may also provide a scaffold for subjacent cornified envelope formation, evidenced by restoration of cornified envelopes after CLE rescue. We provide multiple lines of evidence that the CLE originates as lamellar body-limiting membranes fuse with the plasma membrane: (i) ABCA12 patients and Abca12-/- mice display normal CLEs; (ii) CLEs are normal in Netherton syndrome, despite destruction of secreted LB contents; (iii) CLEs are absent in VSP33B-negative patients; (iv) limiting membranes of lamellar bodies are defective in lipid-synthetic autosomal recessive congenital ichthyoses; and (v) lipoxygenases, lipase activity, and LIPN co-localize within putative lamellar bodies.
Copyright © 2018 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
CONFLICT OF INTEREST
The authors state no conflict of interest.
Figures
References
-
- Akiyama M Corneocyte lipid envelope (CLE), the key structure for skin barrier function and ichthyosis pathogenesis. J Dermatol Sci 2017;88: 3–9. - PubMed
-
- Behne M, Uchida Y, Seki T, de Montellano PO, Elias PM, Holleran WM. Omega-hydroxyceramides are required for corneocyte lipid envelope (CLE) formation and normal epidermal permeability barrier function. J Invest Dermatol 2000;114:185–92. - PubMed
-
- Brash AR. Lipoxygenases: occurrence, functions, catalysis, and acquisition of substrate. J Biol Chem 1999;274:23679–82. - PubMed
-
- Brash AR, Yu Z, Boeglin WE, Schneider C. The hepoxilin connection in the epidermis. FEBS J 2007;274:3494–502. - PubMed
-
- Candi E, Schmidt R, Melino G. The cornified envelope: a model of cell death in the skin. Nat Rev Mol Cell Biol 2005;6:328–40. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
