

Hypothalamic Neurons that Regulate Feeding Can Influence Sleep/Wake States Based on Homeostatic Need
- PMID: 30471995
- PMCID: PMC6279563
- DOI: 10.1016/j.cub.2018.09.055
Hypothalamic Neurons that Regulate Feeding Can Influence Sleep/Wake States Based on Homeostatic Need
Abstract
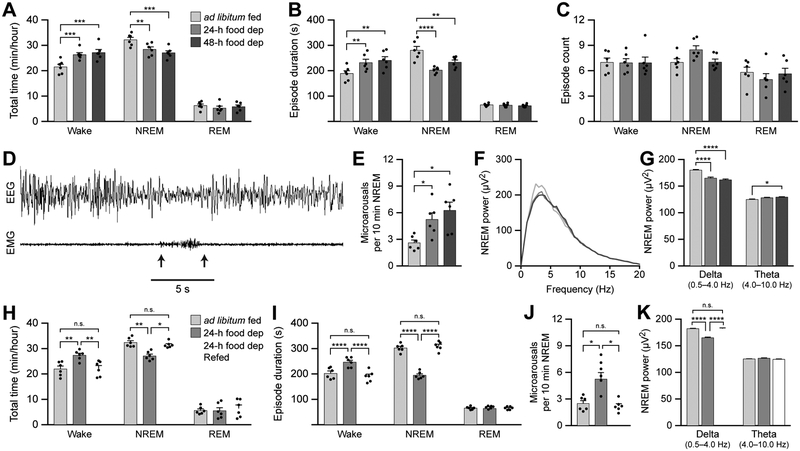
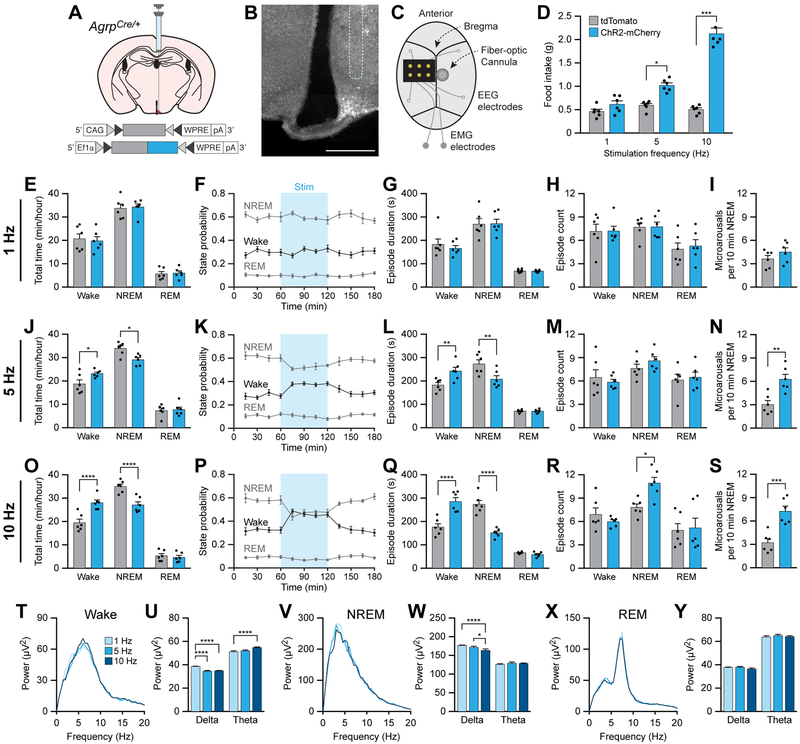
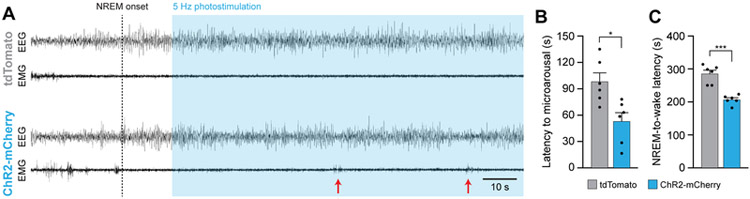
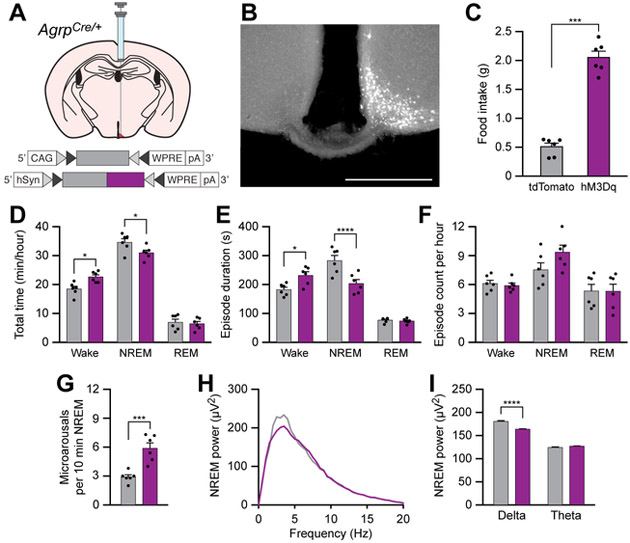
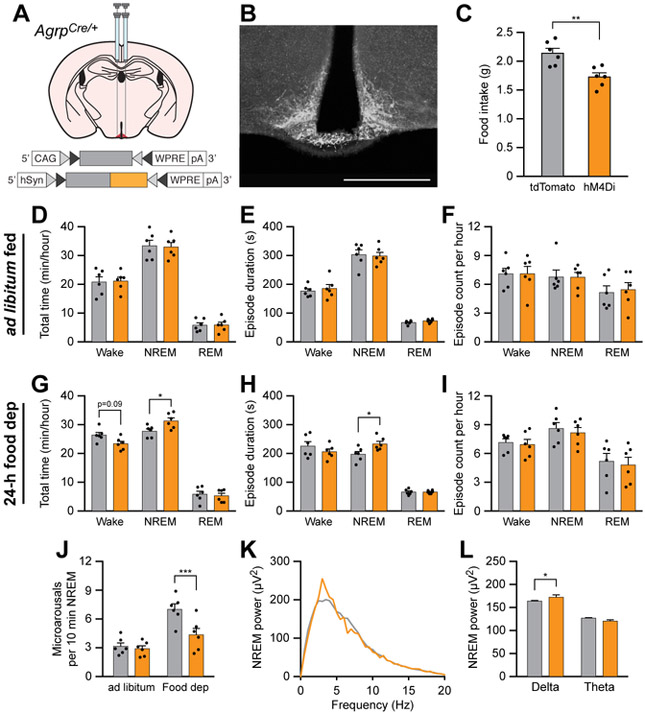
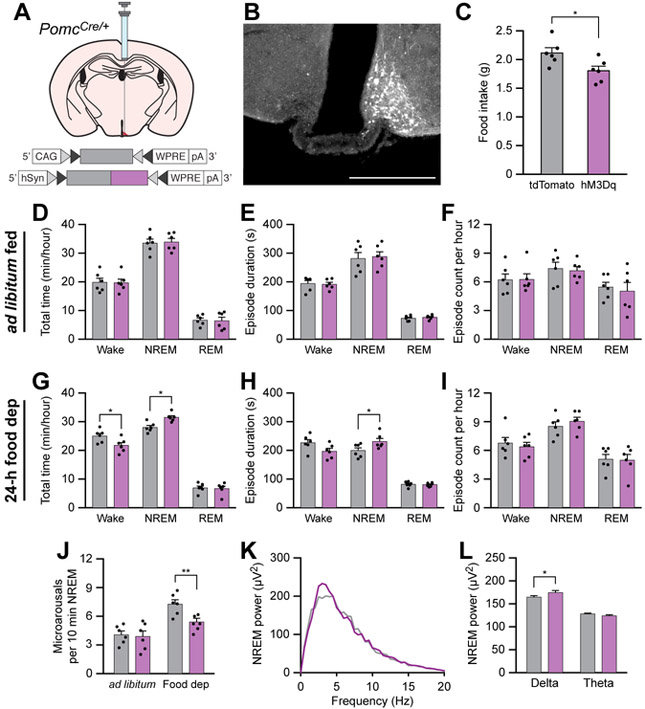
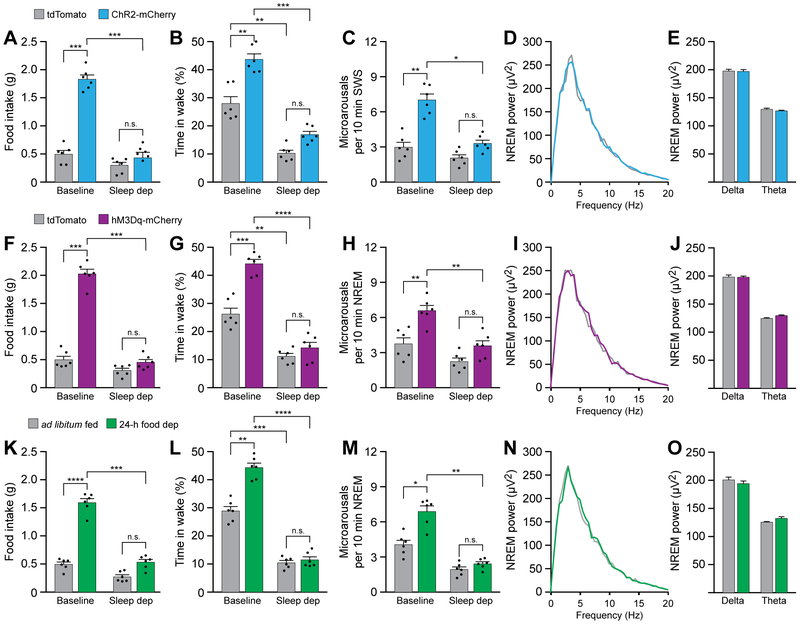
Eating and sleeping represent two mutually exclusive behaviors that satisfy distinct homeostatic needs. Because an animal cannot eat and sleep at the same time, brain systems that regulate energy homeostasis are likely to influence sleep/wake behavior. Indeed, previous studies indicate that animals adjust sleep cycles around periods of food need and availability. Furthermore, hormones that affect energy homeostasis also affect sleep/wake states: the orexigenic hormone ghrelin promotes wakefulness, and the anorexigenic hormones leptin and insulin increase the duration of slow-wave sleep. However, whether neural populations that regulate feeding can influence sleep/wake states is unknown. The hypothalamic arcuate nucleus contains two neuronal populations that exert opposing effects on energy homeostasis: agouti-related protein (AgRP)-expressing neurons detect caloric need and orchestrate food-seeking behavior, whereas activity in pro-opiomelanocortin (POMC)-expressing neurons induces satiety. We tested the hypotheses that AgRP neurons affect sleep homeostasis by promoting states of wakefulness, whereas POMC neurons promote states of sleep. Indeed, optogenetic or chemogenetic stimulation of AgRP neurons in mice promoted wakefulness while decreasing the quantity and integrity of sleep. Inhibition of AgRP neurons rescued sleep integrity in food-deprived mice, highlighting the physiological importance of AgRP neuron activity for the suppression of sleep by hunger. Conversely, stimulation of POMC neurons promoted sleep states and decreased sleep fragmentation in food-deprived mice. Interestingly, we also found that sleep deprivation attenuated the effects of AgRP neuron activity on food intake and wakefulness. These results indicate that homeostatic feeding neurons can hierarchically affect behavioral outcomes, depending on homeostatic need.
Keywords: AgRP; POMC; agouti-related protein; appetite; chemogenetics; homeostasis; optogenetics; pro-opiomelanocortin; sleep.
Copyright © 2018 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
