

Cell cycle-dependent and -independent telomere shortening accompanies murine brain aging
- PMID: 30472697
- PMCID: PMC6286833
- DOI: 10.18632/aging.101655
Cell cycle-dependent and -independent telomere shortening accompanies murine brain aging
Abstract
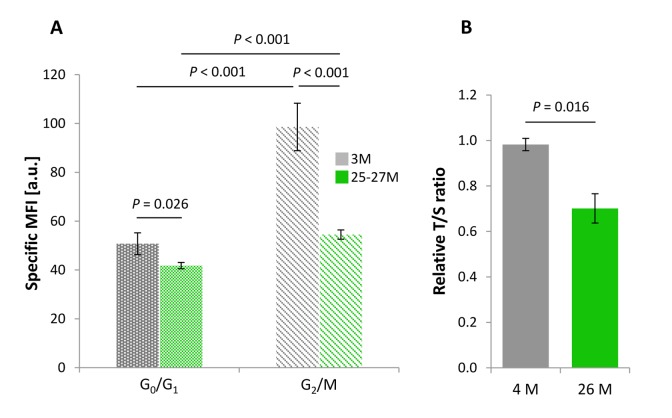
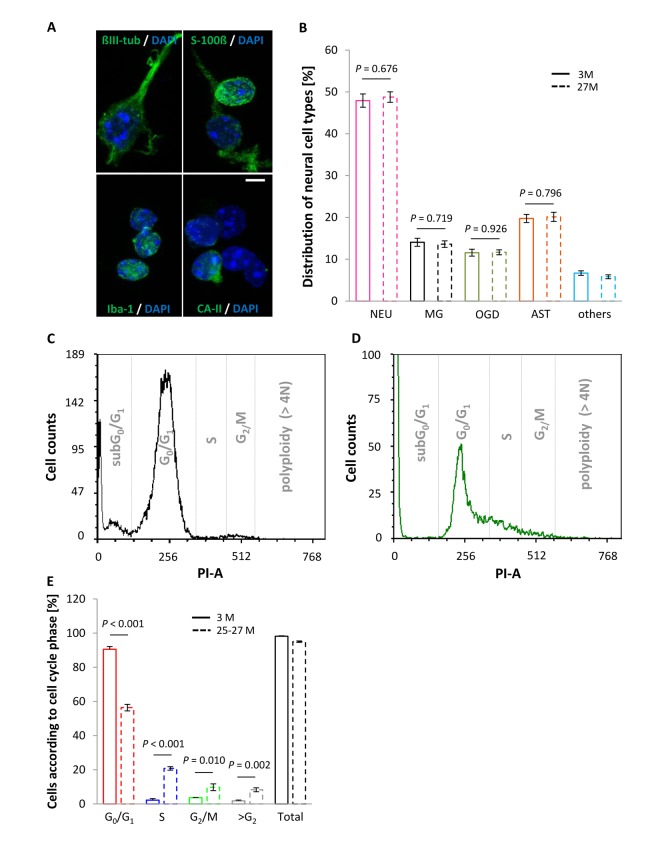
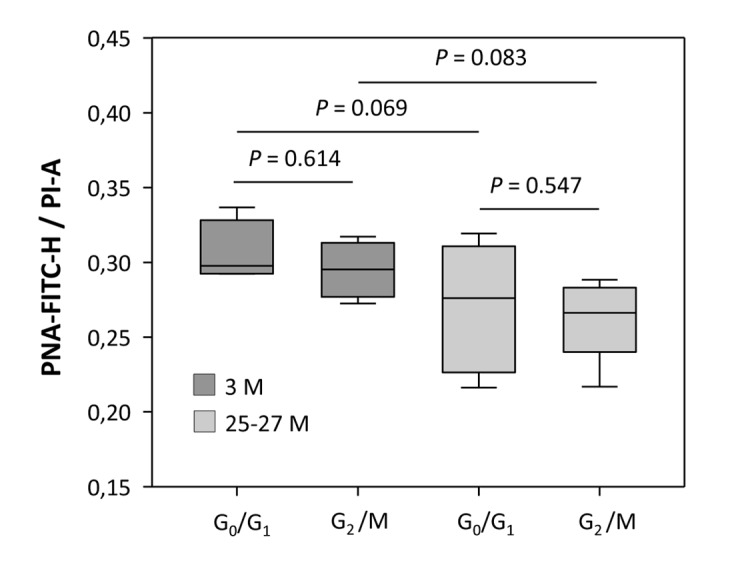
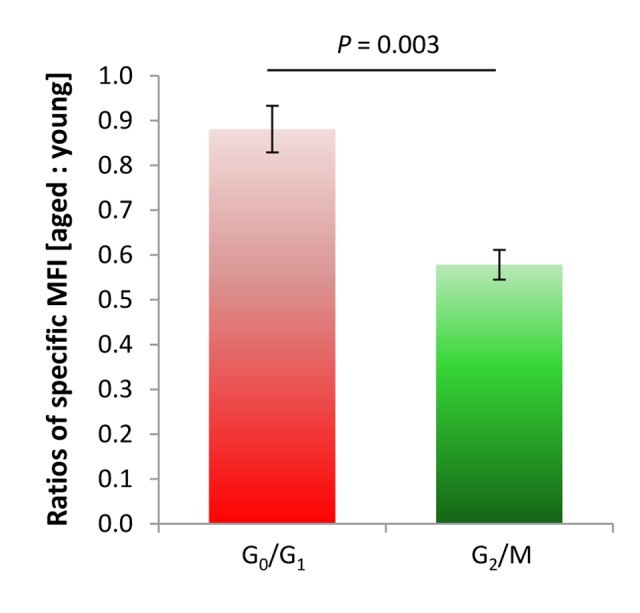
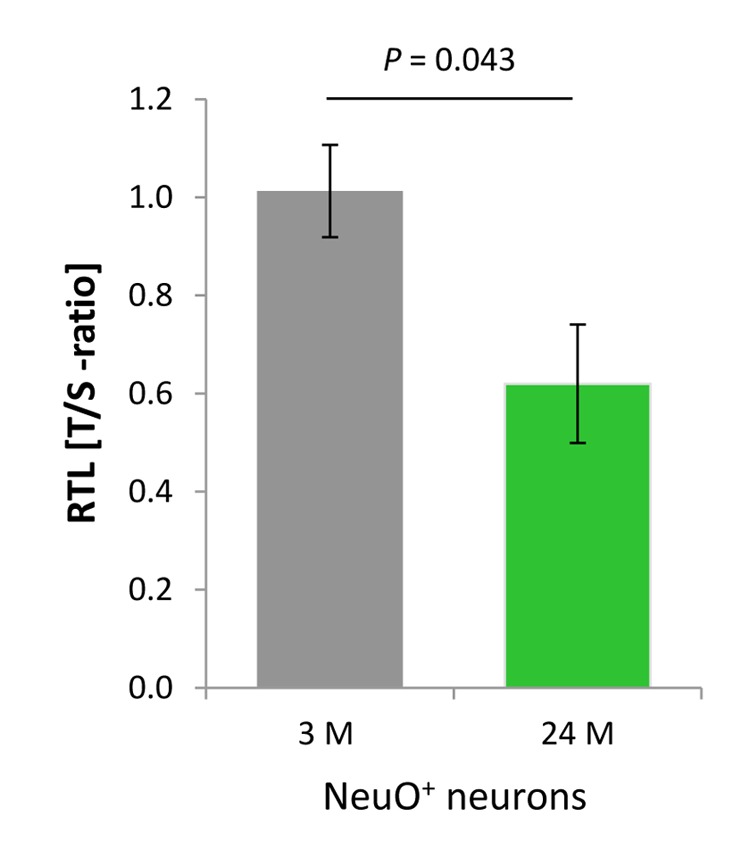
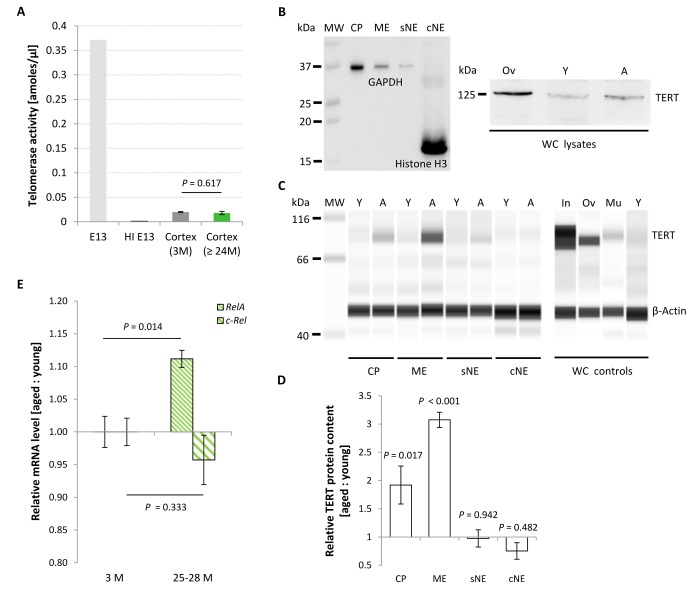
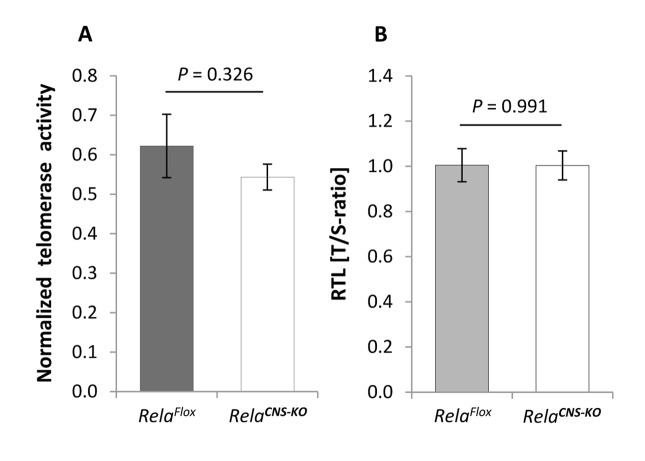
Replication-based telomere shortening during lifetime is species- and tissue-specific, however, its impact on healthy aging is unclear. In particular, the contribution of telomere truncation to the aging process of the CNS, where replicative senescence alone fails to explain organ aging due to low to absent mitotic activity of intrinsic populations, is undefined. Here, we assessed changes in relative telomere length in non-replicative and replicative neural brain populations and telomerase activity as a function of aging in C57BL/6 mice. Telomeres in neural cells and sub-selected neurons shortened with aging in a cell cycle-dependent and -independent manner, with preponderance in replicative moieties, implying that proliferation accelerates, but is not prerequisite for telomere shortening. Consistent with this telomere erosion, telomerase activity and nuclear TERT protein were not induced with aging. Knockdown of the Rela subunit of NF-κB, which controls both telomerase enzyme and subcellular TERT protein allocation, did also not influence telomerase activity or telomere length, in spite of its naive up-regulation selectively under aging conditions. We conclude that telomere instability is intrinsic to physiological brain aging beyond cell replication, and appears to occur independently of a functional interplay with NF-κB, but rather as a failure to induce or relocate telomerase.
Keywords: RelA subunit of nuclear factor kappa B; brain aging; cell cycle; cellular senescence; telomerase reverse transcriptase; telomere length.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
